
Abstract
Savanna rangelands provide diverse communities across southern Africa with livestock and wildlife-based livelihoods, as well as extensive ecosystem services. Historical usage patterns, however, are increasingly challenged by widespread degradation. While regional- and local-scale policy initiatives have attempted to minimize damage and increase the sustainability of savanna rangelands, poverty, land tenure and shifting climate conditions all exacerbate ongoing degradation. Here, we detail the environmental and political setting of southern African savanna rangelands, causes and implications of rangeland degradation, and discuss possible strategies toward improved regional ecosystem management. We present recent knowledge on how degradation by bush encroachment influences biodiversity and biodiversity-mediated ecosystem functioning of semiarid savanna rangelands with the aim of improving rangeland management strategies. Improved rangeland management requires a broad approach which integrates both socioeconomic and ecological frameworks, built upon improved understanding of the strong couplings between flora, fauna, water and land-management strategies.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
1 Biophysical Features of African Savanna Rangelands
Southern Africa’s vegetation is dominated by tropical savannas (Fig. 15.1) which can be distinguished from other vegetation biomes by their codominance of trees and C4 grasses (see Chap. 2). Within the southern African savanna, there is a vast variety of types, such as the different ecoregions as defined by Olson et al. (2001) (Fig. 15.1). They are home to diverse endemic floras and faunas including the earth’s greatest diversity of ungulates. Most of the natural land area of southern Africa can be considered as rangeland (Ellis and Ramankutty 2008), i.e., natural ecosystem habitats managed for grazing livestock and wildlife (Allen et al. 2011).


Topographic Overview of southern Africa based on ETOPO1 data (a). Mean annual rainfall is derived from CHIRPS data and averaged from 1980 to 2020 (Funk et al. 2015). Red line indicates 200 mm/yr isohyet, and the blue line is 400 mm/yr. (b) Land surface temperature is derived from MODIS data 2000–2021 https://lpdaac.usgs.gov/products/mod11a1v006/
Climate
A climatic feature of all southern African savannas, which occur over a wide rainfall gradient from approximately 250 mm to 1800 mm mean annual precipitation (MAP), is almost exclusive summer rainfall with long dry winters (see Fig. 15.2, Huntley 1982; Scholes and Archer 1997). Interannual variability in rainfall, with the most arid areas having the greatest coefficient of variation in rainfall, means that years of both intense drought and near flooding are common (see Chap. 7). The drought and above average rainfall are linked to El Niño–Southern Oscillation cycles and as such periods of either above or below average rainfall are the norm. This has profound impacts on the carrying capacity for livestock and wildlife with animal numbers increasing during rainy periods but with die-offs occurring during droughts (Shackleton 1993).
Summers tend to be hot to very hot, with potentially cool nights (Scholes 2004). Frost occurs in some areas, but the occurrence of heavy frost is a key determinant of the interface between savanna and the true South African grasslands (Fig. 15.1) in moister regions (Ellery et al. 1991). Large portions of south-western savannas have highly variable annual temperature cycles (Fig. 15.3).

Annual temperature seasonality. (a) Long-term (2001–2021) average annual temperature amplitude (e.g., warmest to coldest month). (b) long-term coldest monthly average temperature and (c) warmest monthly average temperature. Data derived from MODIS (https://lpdaac.usgs.gov/products/mod11a1v006/)

Satellite-based Normalized Differential Vegetation Index (NDVI) derived from MODIS data (https://lpdaac.usgs.gov/products/mod11a1v006/). Sixteen-day measurements were averaged from 2000 to 2021 and their mean (a) and standard deviation (b) are shown. Note the low vegetation cover and low standard deviation in northern Namibia. Savanna regions are partly associated with slightly higher standard deviation, suggesting seasonal changes (light red colors in b)
Vegetation and Soil
Due to the high variation in climate and geology, there is a large degree of variation in vegetation types and vegetation amount over the climatic and geological range of the savannas (Figs. 15.1 and 15.4). Changes in geology can be clearly discerned at a regional scale; texture and nutrient status of soils have a profound influence at a local scale. The position on the slope, i.e., the catena position, leads to distinctive changes in vegetation characteristics (see Chap. 2). For instance, on the granite soils of the Kruger National Park, top of slopes are sandy, dystrophic, leached soils and are dominated by broadleaved trees and unpalatable grasses. In the valley at the bottom of slopes, the soil has a finer texture and higher nutrient status and is dominated by fine leaved trees and palatable grasses. A mid-slope hydromorphic grassland fringed by Terminalia species is also common (Scholes 2004).
Scientists differ on how the savannas should be subcategorized. Huntley (1982) points out a clear distinction between what he terms moist/dystrophic savannas (Miombo) and arid/eutrophic savannas (bushveld). In addition, the vast hot, low-lying areas are covered in near monospecific stands of the mopane tree (Colophospermum mopane), and though part of Huntley’s arid savanna is sufficiently different for Olson et al. (2001) to include them as a unique bioregion. Key differences between these three savanna types are given in Table 15.1. Mukwada (2018) suggests that the arid savanna can be broken into five types, moist/mesic dystrophic savanna, dry eutrophic savannas, dry miombo woodlands, mopane woodlands, teak woodlands and Terminalia-Combretum and Acacia woodlands. He, however, makes no attempt to map the distribution of these woodlands. Olson et al. (2001) identify nine savanna types (see Fig. 15.1). Savanna vegetation is often mapped to an even finer detail within individual countries; for instance, in South Africa, Mucina and Rutherford (2006) identify 87 distinct savanna vegetation types grouped into 6 bioregions.
The woody cover of savanna varies from 0% in hydromorphic grasslands through to about 80% in areas of over 600 mm MAP. Above this, the canopy fully closes and the grass layer is lost. Savanna systems persist at tree densities below those that could be expected from rainfall (Sankaran et al. 2005). Many ecologists consider them to be disequilibrium systems where disturbances such as fire, herbivory, droughts and floods are key to their functioning (Scholes 2004). Therefore, any change in herbivory and fire can shift a savanna to either an increased or decreased tree or bush density (see Sect. 15.4.1).
2 Land Tenure and Grazing Systems
The management and use of the savanna rangeland is closely tied to the tenure arrangements, with three dominant tenure patterns being found in all the southern African countries, with country specific nuances. These are commercial tenure, either on freehold or leasehold land, communal tenure and conservation areas on state land (Table 15.2). Early European colonialists tended to establish large commercial farms/ranches that were initially managed almost exclusively for livestock production. These farms tended to be stocked at stocking rates considered to optimize economic return. Production of livestock was almost exclusively for marketing purposes and over time, complex management strategies were developed based on stocking rates and grazing rotation. Individual farmers used different stocking rates and rotations, this despite legislated maximum stocking rates in most countries. Initially wildlife was seen as competing with grazing for livestock and commercial farmers actively prevented wildlife from their land (ABSA 2003). A change in wildlife tenure legislation during the 1970s effectively changed wildlife from being royal game to a commodity which commercial farmers could own and financially exploit (Carruthers 2008). This resulted in a huge conversion of livestock land to mixed livestock and wildlife or simply wildlife. ABSA (2003) found that over 5000 game ranches and 4000 mixed game and livestock farms were developed in South Africa alone. Low value cattle land, such as that adjacent to the Kruger National Park in South Africa, is now some of the most valuable savanna rangeland and being run as exclusive private wildlife destinations. Similar trends to wildlife based land use are found in all the southern African countries.
Communal management of rangelands has historically been for subsistence use of livestock and is very common in much of South Africa. The rangeland is managed as a commonage and is typically stocked at ecological carrying capacity. In some cases, livestock is kept for cultural and investment purposes and offtake for sale is only when cash is desperately needed. In such cases, the driving force in management is to maximize numbers rather than turnover or quality. In Botswana, there is a higher degree of commercial cattle production on the communal rangelands with individual farmers with large commercial herds drilling boreholes to open up new exclusive grazing areas.
There has been, however, a trend to what is termed Community Based Natural Resource Management (CBNRM) in all countries. This started with the CAMPFIRE program in Zimbabwe, but it is probably Botswana and Namibia where this is most developed (Bond 2001). In Botswana, areas that were previously termed Wildlife Management Areas have been converted into Community Management Areas. Many of these areas are now managed by communities for ecotourism or hunting, with wildlife rather than livestock. In Namibia, communal conservancies also opt for wildlife management to gain income from tourism to reduce poverty of communal households. Furthermore, wildlife tourism is promoted as a sustainable land use option and an alternative to livestock farming (Berzborn and Solich 2013). Beyond CBNRM programs, the governments try to correct colonial injustices with land reforms and resettlement programs but with different paths. On one side, the fast track and highly conflictive land reform in Zimbabwe and on the other side the slow resettlement program in Namibia are two extremes of land reform in southern Africa. Land reform is directed mainly to the redistribution of freehold land of white farmers, but also of state land (Breytenbach 2004). CBNRM and land reform contribute to more equity in land tenure and conservation, but until today, it was not possible to turn around colonial injustices. Furthermore, both development pathways generate new borders, conflicts and uncertainties.
Southern African savannas have exceptionally high levels of formal state protection as conservation areas, but the level of actual protection varies across the countries (Table 15.2). Moreover, management in wildlife conservation areas has evolved over time. In the past, there was sometimes extensive management intervention in terms of fire management, elephant density controls or changing the balance of predator and prey species and the provision of artificial watering points. The more recent trend, particularly in the larger reserves, is to now allow systems to regulate naturally with fewer management interventions and a reduction in artificial watering points.
3 Savanna Rangelands as Source of Food, Fodder and Valuable Ecosystem Services
As per their definition, one of the main uses of rangelands is to produce livestock and increasingly wildlife. In addition to this, the savanna rangelands of southern Africa provide many additional ecosystem services critical to maintaining both local livelihoods (Shackleton and Shackleton 2004) as well as being an important global carbon sink (Scholes 2004). The benefits derived from the rangelands differ substantially by the tenure system and management objectives (Table 15.2). Especially in communal areas, rangelands provide the greatest diversity in additional ecosystem services. Shackleton and Shackleton (2004) identify eight categories of direct benefits: fuelwood, construction material, wild foods, medicine, household implements, fodder, fibers and cash sales, as well as four indirect benefits: spiritual, cultural, indigenous knowledge and ecological services accruing to community’s resident within the savanna rangelands. A multitude of studies suggests that the provisioning of fuelwood is often the single biggest value of the savanna rangelands to local communities (e.g., Dovie et al. 2004). Most rural communities are almost fully dependent on fuelwood for household cooking energy. The wood use varies widely, depending on household size and dead wood availability, with Dovie et al. (2004) estimating approximately 700 kg/capita/year usage. Use of charcoal is not that common in rural areas, but is a major fuel source in many cities, with the communal areas deriving substantive income from the sale of charcoal (Baumert et al. 2016). Charcoal trade differs substantially across the subregion. South Africa, Botswana, eSwatini and Zimbabwe having relatively low or no charcoal dependency, while Mozambique, Zambia, Malawi and Tanzania having urban centers almost totally dependent on charcoal as the fuel for low income households. For instance, the town of Maputo has shown an increase from most households preferring firewood (60%) to charcoal (17%) in 1990 to 87% of informal households using charcoal in 2004 and 96% using charcoal by 2016 (Mudombi et al. 2018). Estimates from Tanzania suggest that rural income from charcoal production exceeds the value of rural subsistence agriculture (Luoga et al. 2000), though harvesting for charcoal is often done by external contractors, with limited financial benefit accruing to the local residents (Baumert et al. 2016). Namibia is a major producer of charcoal from eradicating bush encroachment, much of this is exported (Shikangalah and Mapani 2020).
In communal areas, rangelands provide a critical safety net during times of need such as droughts (Shackleton and Shackleton 2004). Although fruits, nuts, insects and mushrooms provide an important nutritional supplement during droughts, these are indispensable to household wellbeing. In addition, the poor in the village often use woodland products for applications such as brooms, or even make and sell these and other crafts for a cash income (Shackleton et al. 2010).
Cattle and other livestock have important cultural value throughout the subregion (Shackleton and Shackleton 2004). In addition to provision of fodder, the savanna rangeland also provides material for the building of cattle kraals (corrals), and indeed for the construction of traditional homesteads which use both poles for the structure and grass for thatching (Dovie et al. 2004).
Medicinal plants from the rangelands are also very important and valuable from both a cultural, spiritual and medicinal perspective. Bulbs, bark and leaves are harvested for a wide variety of treatments (Mander 1998).
In commercial areas, the use of rangeland has traditionally focused on subsistence-based livestock production, with limited additional benefits to the land-owner. A trend in all southern countries has been a move from livestock toward mixed livestock and wildlife or to total wildlife.
In conservation areas, the key benefit from the rangeland is biodiversity preservation. However, in these and all other rangeland areas, there is also growing realization of the important role of the savanna rangelands in terms of being a globally important carbon sink (see Chap. 17).
4 Indicators and Drivers of Degradation in African Savanna Rangelands
Rangeland degradation is a growing threat to many savanna regions in southern Africa. Degradation is expressed through lowered groundwater tables and reduced water quality, soil erosion and decreased soil fertility, and both loss of—and changes in—plant species density, diversity and palatability as well as bush encroachment. Both climatic (e.g., changes in precipitation, CO2 levels) and anthropogenic (e.g., overgrazing, fire suppression and the socioecological framework) drivers contribute to rangeland degradation through overlapping and often synergistic processes.
4.1 Climate, Atmospheric CO2, Overgrazing and Fire Suppression
The consensus explanation for savanna degradation is that overgrazing reduces the grass layer, which in turn increases the opportunities for bush establishment by lower fuel loads for natural fires and reduced grass competition with emerging bush seedlings (e.g., Walter 1939; Archer et al. 1995; Higgins et al. 2000; Ward 2005). When grasses disappear, bare-soil patches become increasingly prevalent, and are highly vulnerable to soil erosion by wind and water.
Both browsing wildlife and natural fires play an important role in maintaining the coexistence of grasses and bushes in healthy savanna ecosystems. The replacement of indigenous browsers and grazers by domestic livestock and the artificial prevention of hot fires are key anthropogenic drivers of intensified rangeland degradation. Indigenous browsers in general suppress bush establishment (Trollope and Dondofema 2003); elephants even uproot mature trees (Dahlberg 2000). Frequent fires in moist savannas limit bush density by killing bush saplings and even large trees if their fire-resistant bark has been stripped by browsers (Yeaton 1988). In more arid savannas, where fires are less frequent, fires often coincide with a series of above-average rainfall seasons. High rainfall generally encourages abundant grass biomass that is then available as fuel for fires (Joubert et al. 2008). It is exactly those wet years where soil moisture conditions are also optimal for bush sapling establishment (Joubert et al. 2008; Lohmann et al. 2014).
Under changing climate conditions—particularly increased rainfall variability and extended droughts—the rate and scale of rangeland degradation is expected to increase; however, site-specific soil, vegetation and land use characteristics make generalization difficult (e.g., Ward 2005; Bond and Midgley 2012; Stevens et al. 2016; Archer et al. 2017). Evidence increasingly suggests that higher atmospheric CO2 will play an additional role in driving rangeland degradation by reducing the advantage in water use efficiency of C4 grasses compared to C3 bushes and trees (Archer et al. 1995; Bond and Midgley 2012; Stevens et al. 2016).
4.2 Socioecological Framework and Policy
The relation between drivers of unsustainable land use (overgrazing and fire suppression) and their sociopolitical and socioeconomic context is rooted in colonialism and apartheid. Colonial expansion was tantamount with grabbing of indigenous lands by white settlers, mining- and farming companies (52% in Namibia in 1990, 87% in South Africa in 1936) (Potts 2012). The establishment of nature reserves for wildlife protection by the colonial authorities exacerbated the loss of land for indigenous communities (Berzborn and Solich 2013). The subsequent territorial encapsulation of indigenous people on remaining lands led to overpopulation in comparison to the white farming areas, which were characterized by large farm sizes (Schnegg et al. 2013). Furthermore, the communal lands in Namibia and South Africa often had worse climatic, geomorphological and soil conditions (Hoffman and Todd 2000; Menestrey Schwieger and Mbidzo 2020). These conditions together with overpopulation led to overstocking in communal areas, despite each communal farmer keeping only few livestock. The villagization, induced by colonial authorities and the apartheid regime, in combination with kraaling and cultivation near the homestead to be able to protect livestock and crops worsened the situation. It led to overgrazing and trampling within and near the settlements. Illegal fencing of communal land by those who can afford fencing materials increases grazing pressure in the remaining commonage by effectively excluding other residents from using the fenced off area. Although it is illegal, it has been observed in communal areas of Namibia. However, fencing by a community could be viewed as a strategy to deal with drought conditions by protecting grazing resources for the future (Kashululu and Paul 2020). In the region of today’s Namibia, the combat against animal diseases led to further territorial encapsulation of black smallholders. The veterinary cordon fence (VCF) dividing the northern areas from the rest of the country was supposed to protect the farming area south of the VCF from animal diseases such as foot and mouth disease (FMD) and to secure export markets. Since the mid-1960s, outbreaks have been limited to the communal areas north of the VCF (see Chap. 18; Schneider 2012). While the farming area south of the VCF, where nearly all white farmers and also communal areas are located, has been well protected against FMD, the new borders and fences represented a conspicuous obstacle to pastoral mobility.
In what is now Namibia and South Africa, before the advent of colonialism, indigenous communities used pastoral grazing systems similar to modern rotational grazing and veld fires to control bush encroachment and to stimulate grass growth (Beinart 2003; Hoffman 2014; Rohde and Hoffman 2012; Menestrey Schwieger and Mbidzo 2020). Colonial authorities in South West African Hereroland and other reserves and the Native Trust in South Africa forcibly intervened into the practices of black smallholders restricting livestock movement, dictating maximum stocking numbers, fencing-off land and establishing grazing or stock fees. As a result, the communal farmers’ mobility, on which their traditional farming practices were based upon, was further reduced, which subsequently intensified overgrazing and trampling. (Delius and Schirmer 2000; Menestrey Schwieger and Mbidzo 2020).
Furthermore, state authorities banned and discouraged the practice of veld fires for freehold as well as communal farmers in the late nineteenth and early twentieth centuries. They feared that fires would damage vegetation and get out of control, even though the beneficial properties were known (Beinart 2003; Rohde and Hoffman 2012; Hoffman 2014; Humphrey et al. 2021). By disrupting traditional pastoralist activities and intentionally giving scarce resources to indigenous communities, colonial authorities and the apartheid regime enforced the diversification of pastoralist activities of communal farmers, which are still practiced today. Absentee farmers work outside of communal areas and use their wages to buy cattle and to increase their herds, which are herded by family members or paid herders. Herd sizes of absentee farmers tend to depend more on their wages than on environmental considerations (Schnegg et al. 2013).
The colonial and apartheid state in today’s Namibia and South Africa established several assistance programs for water supply infrastructure, livestock vaccination, new livestock breeds, farming techniques and the drought relief programs. The governmental assistance enabled livestock farming beyond natural limits and avoiding destocking in the face of drought especially for freehold farmers showing favorable treatment of white land users (Lange et al. 1998; Delius and Schirmer 2000; Bollig 2013; Kreike 2009; Menestrey Schwieger and Mbidzo 2020). Summing up, the disruption of pastoralist practices of communal farmers as well as the favorable government assistance for freehold farmers led to overstocking and in the end land degradation.
5 Degradation of African Savanna Rangelands
5.1 Bush Encroachment
Bush encroachment has been defined as the increase in the density of woody vegetation in grassland and savanna ecosystems (e.g., De Klerk 2004; Smit 2004; Ward 2005; O’Connor et al. 2014; Archer et al. 2017). It occurs across all southern African savannas (Fig. 15.5), with increasing severity along a moisture gradient (O’Connor et al. 2014).

Changes in nonforest and nonanthropogenic landcovers. (a) MODIS landcover classification 2019 grouped by functional type. Note the preponderance of shrublands and grasslands in much of southern Africa. (b) Changes in landcover (2001–2019). Blue areas show a move toward more woody (e.g., woody savanna, shrublands) landcover, brown areas show a move toward more grassy (e.g., grasslands, open savanna) landcover. Grey areas show no change in landcover type, as well as anthropogenic or other natural (e.g., forest) landcovers. More land area has shifted toward grasslands; however, the spatial pattern of these changes is complex (https://lpdaac.usgs.gov/products/mod11a1v006/)
Bush encroachment describes a change in savanna vegetation structure and function impacting ecosystem services and ecosystem disservices worldwide (e.g., Li et al. 2016). From a rangeland perspective, it is regarded as a major form of land degradation. It diminishes the availability and quality of forage for grazing animals, with impacts on meat, milk and leather (Shikangalah and Mapani 2020). Bush encroachment can also lead to an increase in certain animal diseases (e.g., Bollig and Osterle 2008). Bush thickets have been blamed for offering hiding spots for predators to ambush livestock (Vehrs and Heller 2017). The impenetrability of thorny thickets increases animal injuries and hinders the accessibility of limited grass forage (Mcleon 1995). A study in Zimbabwe showed that for every percent increase in bush cover grass biomass decreased by 26.5 kg/ha. In the Molopo region in South Africa veld productivity for cattle was suppressed by 970 kg/ha of dry matter resulting in a reduction in grazing capacity from 8.7 to 45 ha per large stock unit (LSU) (Moore and Odendaal 1987). Wigley et al. (2009) reported a 20% decline in grazing capacity in a mesic savanna rangeland in KwaZulu-Natal, South Africa over a 63-year period between 1937 and 2000. In Namibia, where the original carrying capacity (also known as grazing capacity) before bush encroachment was 10 ha/LSU it decreased to 30 ha/LSU (de Klerk 2004; NAU 2010). Due to the reductions in grazing capacity and related consequences, in 2012, the Namibian Meat Board estimated a decline of about 30% to 64% in cattle numbers in commercial farms when compared to 1959 (Demas et al. 2012). This decline was estimated to be equivalent to an economic loss in meat production of N$700–1.6 billion (US$ 94.8 million–US$ 217 million) per annum (NAU 2010; Trede and Patt 2015). For Zimbabwe, woody encroachment caused a loss of £16 million (US$74 million) in beef production already in 1948 (Hattingh 1952). This is all worrisome and can lead to significant declines in food security, such as in Namibia, where livestock production (mainly beef production) is estimated to contribute up to 75% to the total agricultural output (Shikangalah and Mapani 2020).
Dense bush also negatively impacts important cultural services especially tourism by obstructing game viewing (Demas et al. 2012) and by decreasing densities of certain wild animals (e.g., Wigley et al. 2009; Demas et al. 2012). In communal areas, the cultural standing as well as the royal value is compromised where bush encroachment prevents cattle production, because cattle are a powerful status symbol in many southern African cultures (Reed et al. 2015).
Despite the apparent and predominantly negative impacts of bush encroachment on cultural services, fodder and animal production, recent studies increasingly show positive effects of structural changes for, e.g., soil properties, biodiversity and associated ecosystem functioning (see Sects. 15.5.3 and 15.5.4). Also, the relatively new consciousness is that Carbon (C) sequestration by woody plants and changed soil organic carbon stocks are potentially an important regulating service of bush encroachment, and can play a vital role in reducing C emissions into the atmosphere globally (Tallis and Kareiva 2007). Although global patterns are not consistent, a study in South Africa showed that soil C content and soil C stocks were on average 148% and 117% greater, respectively, in bush-encroached compared to open grasslands (Dlamini et al. 2019, see also Chap. 17).
5.2 Soil Erosion, Soil Nutrient and Soil Moisture Decline
High stocking rates of livestock or wildlife can decrease soil water by two processes: (i) trampling can destroy soil porosity, which decreases infiltration and (ii) grazing decreases plant cover critical for soil organic matter, which is important for water storage (water holding capacity) (McNaughton et al. 1988). High stocking rates reducing the density of vegetation cover and size of vegetation patches (overgrazing) also increase soil erosion, thus leading to an overall loss of water and nutrient resources from the ecosystem (Rietkerk and van de Koppel 1997). Degradation of soil resources will largely determine primary production of vegetation (fodder) also in the long-term.
It has been argued, however, that the increase in thick dense bush and the spread of the unpalatable grasses, which is normally viewed as degradation of savanna rangelands, may actually help controlling soil erosion (e.g., Shikangalah and Mapani 2020). Aboveground plant parts covering the ground decrease the erosive impacts of raindrops and high winds. The root system anchoring the soil offers stability to the soil structure. Moreover, bush encroachment mainly by legumes that fix nitrogen increases nitrogen mineralization. Greater soil fertility with bush cover was observed in many African savanna rangelands (Belsky 1994; Smit 2004; Sitters et al. 2013; Dlamini et al. 2019; Sandhage-Hofmann et al. 2020; Mogashoa et al. 2021).
While nitrogen-fixing legumes may indeed increase nutrients in the soil and can improve the soil microclimate by shade (Metzger et al. 2014), they possibly deplete soil moisture by high evapotranspiration and lowered infiltration (Archer et al. 2017). Bush encroachment alters soil moisture availability (Geissler et al. 2019a) and will largely determine both the recovery potential of the remaining herbaceous vegetation and the establishment of new bush seedlings leading to further bush encroachment.
5.3 Decline in Water Quality and Groundwater Recharge
Although often less obvious, a decreased water quality (i.e., surface and subsurface water) is another important sign of rangeland degradation. A water quality parameter of concern in many catchments of this world is nitrate (e.g., Weitzman et al. 2021), since the very complex biogeochemical nitrogen cycle can be influenced in various ways. Regarding rangeland degradation, a shift in species composition and/or biomass production, particularly toward nitrogen fixing Senegalia species and other legumes, will alter the nitrogen cycle in the unsaturated zone and consequently influence the nitrate concentration in groundwater carried in with infiltration water.
Poor water point management (Fig. 15.6) can further the impacts and amplify groundwater pollution. Moreover, water points are areas of the highest animal excrement deposition. Soil erosion could free nitrate pools, but also lead to a loss of fungi and bacteria that are usually involved during ammonification in the soil zone, an intermediary step of the nitrogen cycle toward nitrate (Hiscock et al. 1991).

Poorly managed water point in an area that is already vulnerable to nitrate contamination by its shallow soil and dominated by Senegalia species. Note: water is leaking from the trough and dissolves nitrate from animal feces that can contaminate groundwater
An example of dynamics in water quality in rangelands is drawn from an area in south-western Etosha National Park, including surrounding conservancies, livestock farms and private game reserves using water quality index (WQI). Among other easy measurable parameters, this index includes nitrate concentration. Comparison of historical (pre-2000) and recent (2020) data of 68 boreholes shows a reduction in water quality at 40% of the boreholes. The number of boreholes with deteriorated water quality in livestock farms and private game reserves was at least two times higher than in conservancies and national park. In contrast, the number of boreholes with improved water quality was at least two times lower. This shows that signs of rangeland degradation in terms of water quality are more pronounced in livestock and private game reserves than in conservancies and national parks.
The effect of bush encroachment on groundwater recharge is a topic of controversy since the differences in water consumption between grasses and bushes are still unclear (Scholes and Archer 1997; O’Connor et al. 2014). Nevertheless, water loss in Namibia through bush encroachment is estimated to be around 12 million m3 on 10,000 ha (NAU 2010). Indeed, Groengroeft et al. (2018) found deep drainage about 3-fold lower in the area below the canopy than in intercanopy patches. They explain the decrease in the amount of deep percolating water by lower infiltration and higher evapotranspiration in bush dominated patches and conclude that bush encroachment is likely to reduce groundwater recharge.
5.4 Biodiversity
Bush encroachment is associated with negative, but also neutral and positive consequences for species richness of plants and animals, and biodiversity-associated ecosystem functioning as part of the degradation process in African savanna rangelands (e.g., Chown 2010; Eldridge et al. 2011). A decline in ecosystem functioning has serious consequences for ecosystem services and the regulatory processes of the systems, which can further exacerbate degradation. A higher biodiversity increases both, the multifunctionality and the resilience of an ecosystem. The so-called “Insurance Hypothesis” assumes that higher species richness increases the probability that functionally redundant species buffer a specific function against environmental fluctuation or future threats. (Buisson et al. 2019).
5.4.1 Plant Diversity
The impacts of bush encroachment on plant diversity in savanna rangelands depend on local conditions of climate and soils, in particular on the level of disturbance and resource stress (soil moisture and nutrients, see Sect.15.5.3), the heterogeneity of these resources and viable soil seed banks. Several studies have reported a decline in the diversity of understory vegetation with increasing bush cover (e.g., Scholes and Archer 1997; Angassa 2005; Mogashoa et al. 2021). Other studies found no effect or a decrease in diversity only above a certain bush cover and only for specific functional groups (perennial grasses, forbs) (Dreber et al. 2018). In contrast, Belay et al. (2013) showed that plant diversity increased with bush density.
Bush encroachment also supports the invasion of alien species (e.g., Chromolaena in South Africa, Wigley et al. 2009), although the susceptibility of rangelands varies between the vegetation types. By changing the structure and function of soils, invasive alien plants exacerbate the risk of losing native plant diversity. The Southern African Plant Invaders Atlas (SAPIA) database contains currently records for over 500 invasive plant species in South Africa, Lesotho and eSwatini (Zengeya and Wilson 2020).
5.4.2 Plant Diversity Mediates Soil Moisture, Groundwater Recharge and Primary Production
The hydrological cycle is in various ways interlinked with biodiversity and ecosystem functions. The availability of soil water obviously controls the primary production of vegetation. Less obvious are biodiversity-mediated feedbacks on the water cycle, controlling local climate and ultimately the availability of freshwater as an ecosystem service to the human beneficiaries.
Depending on plant size and morphology, rainfall is differently intercepted by the above-ground parts of plants and either directly returned to the atmosphere via evaporation from the canopy or funneled to the soil below via stemflow and throughfall (Yuan et al. 2016). Water reaching the soil can affect not only the amount of water available to plants but also the amount of groundwater recharge and runoff, controlling soil erosion. However, most dryland plants—particularly those in Africa—are poorly explored. Comparing the impacts of morphologically different African bush encroacher species such as Terminalia sericea, Dichrostachys cinerea, Colophospermum mopane, Senegalia mellifera, Vachellia reficiens and Catophractes alexandri on rainfall interception and subsequent soil moisture is key focus of the recent research program SPACES.
Once water has entered the soil, diverse rooting systems of different plant species can either improve local soil-water availability by increasing the preferential flow along the root systems into deeper soil layers or decrease local soil-water availability by a complete uptake of water via spatial niche partitioning (Lee et al. 2018). Modern stable isotopes analyses of water indicate a sharing of water resources between many species of bushes, trees and grasses in the upper soil of African savanna rangelands (Kulmatiski et al. 2010; Beyer et al. 2018; Geissler et al. 2019b; Uugulu 2022). These results indicate that diverse vegetation has a potentially higher water use efficiency (biomass produced per unit water used) compared to species-poor ecosystems. High water-use efficiency maximizes the relative amount of water kept in the ecosystem; however, groundwater recharge potentially decreases as fewer precipitation events reach below the effective root zone. The positive impact of plant diversity on water use efficiency was confirmed by an ecohydrological simulation study (Irob et al. 2022). The simulation results also showed a positive effect of functional diversity on vegetation cover, especially the cover of perennial herbaceous vegetation. As a result of these changes, the biomass production of sites with functionally diverse vegetation increased, leading to improved fodder production.
5.4.3 Animal Diversity
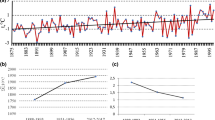
Bush encroachment will strongly impact animal diversity in African savanna rangelands through changes in the structural diversity of vegetation. Structural diversity not only determines the amount and quality of habitats (i.e., niche space), but also the heterogeneity of resources and the interactions between species (e.g., Hering et al. 2019). While an initial increase in bush cover increases structural diversity, severe bush encroachment has the opposite effect (e.g., Blaum et al. 2007a)—it creates homogeneous and structure-poor environments that lead to the loss of habitats and species niches. Thus, animal diversity often shows hump-shaped responses to increasing amounts of bush cover (e.g., Chown 2010). Studies along bush-cover gradients in the Kalahari in South Africa have shown similar responses for insects and spiders (Blaum et al. 2009; Hering et al. 2019) and for small- to medium-sized mammalian carnivores (Blaum et al. 2007a, b). Maximum diversity for all taxonomic groups was found at a mean bush cover of 15% (Fig. 15.7). Similar responses to altered structural diversity linked to bush encroachment have been reported for birds in the Rooipoort Nature Reserve in South Africa (Sirami et al. 2009) and lizards in central Namibia (Meik et al. 2002). Though hump-shaped distributions recur for some taxonomic groups, linear negative effects of bush encroachment were observed for reptiles (Wasiolka and Blaum 2011) and rodents (Blaum et al. 2007c; Fig. 15.7).

The loss of structural diversity due to degradation of savanna vegetation may even cascade up to large herbivores and top predators. In South Africa’s Kruger National Park, the reduction of grass cover induced by an increasing bush layer led to declines in grazing herbivores such as zebras (Equus burchellii) and wildebeests (Connochaetus taurinus), which in turn were replaced by browsing herbivores such as kudus (Tragelaphus strepsiceros) and giraffes (Giraffa camelopardalis) (Smit and Prins 2015). Likely, the changes in herbivore communities caused by structural changes in vegetation negatively affect top predators. Lowered preferred prey densities, poor visibility and restricted movement spaces may reduce their hunting efficiency and alter hunting behavior (e.g., Muntifering et al. 2006).
Significant changes in hunting behavior due to bush encroachment have already been noted for different taxa. For example, the Spotted sand lizard (Pedioplanis l. lineoocellata) is classified as a “sit and wait forager.” That is, it sits and hides in a grass tussock and waits for prey to pass by. In bush dominated habitats with low food availability, the Spotted sand lizard changes its foraging behavior to “actively foraging.” Individuals actively search for prey and travel significantly longer distances, which increases their own predation risk. This behavioral plasticity acts as a buffer mechanism against the negative effects of bush encroachment and allows this species to still occur in heavily bush encroached savannas, albeit at significantly lower densities (Blumröder et al. 2012).
These examples illustrate that consequences of changes in structural diversity caused by bush encroachment may vary considerably across taxonomic groups and that multiscale feedback mechanisms can impact trophic cascades and complex interactions in food webs.
5.4.4 Animal Diversity Mediates Soil Moisture and Soil Nutrient Dynamics
The awareness of animal biodiversity for ecosystem functioning has developed rapidly in the past decades. Early work has focused primarily on indigenous large mammals and has largely overlooked the role of small animals only until recently. Bioturbating animals such as termites, ants, beetles and other invertebrates can alter the micro-soil environment. They modify soil texture, build tunnels (macropores) and transport nutrients into the soil (e.g., termites, dung beetles) that affect a variety of soil functions, such as nutrient accessibility, water holding capacity and water sorption (Rückamp et al. 2012), which in turn impact vegetation (e.g., Traore et al. 2015). Termites for example, promote plant growth directly around their nests, thereby altering the vegetation composition and structure of entire landscapes (Bonachela et al. 2015).
Macropores of all sizes and forms created by termites, ants or beetles that open to the soil surface can strongly improve water infiltration into soils (e.g., Léonard et al. 2004; Brown et al. 2010; Colloff et al. 2010). Precipitation can preferentially flow into deeper soil layers, making it less susceptible to evaporation loss and thereby improving the ecosystems water balance and groundwater recharge (Bargués Tobella et al. 2014). This effect is stronger on soils with higher loam content, compared to sandy soils (Marquart et al. 2020c). Loamy soils with termite burrowing activity can act primarily as runoff interception areas and thereby reduce water loss and soil erosion (Léonard et al. 2004); however, some studies have found that the termite-built sheetings (i.e., water-repellent compacted surfaces) decrease infiltration under dry conditions, while the construction of tunnels increases infiltration under wetter conditions (Cammeraat et al. 2002).
The transport of feces into the soil by dung beetles (Nichols et al. 2008) and the subsurface nests build by meat ants (Iridomyrmex greensladei) (Nkem et al. 2000) enhance nutrient cycling and soil aeration, and can also counteract soil compaction and improve water infiltration compared to surrounding soils.
Growing evidence suggests that soil-burrowing invertebrates in general may enhance the restoration success of degraded rangelands (Colloff et al. 2010; Kaiser et al. 2017). Since, macropore abundance is generally higher under bushes compared to the interspace (Marquart et al. 2020a, b), restoration measures such as area-wide de-bushing should keep single bush patches intact. These patches provide important microhabitats and can preserve founder populations of soil invertebrate communities, hence maintaining their ecosystem function and improving ecosystem restoration efforts.
6 Impacts of Degradation on Human Livelihoods
With ecosystem degradation, the lives and livelihoods of those dependent on ecosystem services become vulnerable and in the case of the poorest, they might even get devastated. According to the Consortium on Ecosystems and Poverty in sub-Saharan Africa (CEPSA), the livelihood of most people living in arid and semiarid African rangelands directly depends on natural resources (Shackleton et al. 2008) through agriculture, fishing and hunting (Barbier and Hochard 2018). The equivalent income share of such products might constitute up to one third of total income of these households. Therefore, declining ecosystem services often (i) lead to a steady erosion of livelihood assets, (ii) increase vulnerability by making people less able to withstand external shocks, (iii) increase the risk of widespread disaster and (iv) might even exacerbate existing conflicts and give rise to new conflicts over access to ecosystem services (Shackleton et al. 2008). In other words, rangeland degradation is not only linked to increased poverty, but rather entails serious consequences for poverty alleviation in general.
One of the most obvious signs of land degradation is declining biomass production. It leads not only to reduced agricultural crop productivity but also to difficulties to support livestock numbers in rangeland systems (Tully et al. 2015) affecting the livelihoods of both pastoral communities and cattle farmers. To manage such lands and support livestock, an increased investment of management costs (i.e., supplementation or bush control) would be necessary (e.g., Lal 2015). In 2016 in Namibia, animal fodder was ranked as the eighth largest imported good, with total fodder imports valuing close to 4 billion Namibian Dollar (NAD), equivalent to the gross value addition of the entire agricultural sector (Honsbein et al. 2017). Such economic investments will, however, only be accessible to those who can afford them and may on top of that be coupled with indebtedness, especially in the case of subsistence and poor farmers (Gomiero 2016).
By the end of the last century, 75% of the population of sub-Saharan Africa depended on subsistence farming (Sanchez et al. 2007). In 2017, an estimate of 4 million households practiced subsistence farming in South Africa (RSA 2017) and in Namibia, today, approximately 48% of rural households depend on it (MET 2020). Most people living in rural areas and depend on agriculture have relatively low purchasing power and are thus rarely able to purchase the required supplementary fodder that livestock needs for a healthy growth and weight gain. Therefore, livestock is undernourished, which in turn implicates an insufficient nutrition and calorie provision for consumers (Schnegg et al. 2013). Moreover, with underweight livestock, potential sales are not as profitable and household capital is confined (Barrett and Bevis 2015). The consequence is a restriction to invest in more sustainable practices, let alone the ability to provide sustenance (i.e., food, health).
Degradation of rangelands can therefore force farmers to abandon their lands and either look for new ones to meet the surviving needs of current and future generations (Gomiero 2016), or look for new opportunities to earn a livelihood elsewhere. Such a situation might also lead to the diversification of pastoralist activities or migration movements. Rangeland degradation can therefore foster urbanization via rural-urban migration (Marchiori et al. 2012). This in turn entails far-reaching consequences, e.g., a loss of local knowledge in rural areas, disintegration of local communities, wage competition in urban areas and as a result migration decisions of urban households. However, given the lack of substantial assets, household members of the poorest seeking additional working opportunities (off-farm or outside of agriculture) tend to seek them locally or migrate only temporarily for short distances (Banerjee and Duflo 2007). This still forces many to exploit natural resources in their surrounding environments to supplement their consumption and income or to support their family members, giving the potential for a downward spiral if no sufficient pathways to improve livelihoods can be offered (see also Barbier and Hochard 2018; Suich et al. 2015).
Besides an increased vulnerability to poverty, land degradation can also dampen economic growth, especially in countries where agriculture and livestock farming is the engine for economic development. Many African economies are very dependent on climate sensitive sectors such as agriculture, forestry and fishery (Diao et al. 2010). Without proper sustainable land management practices to reduce rangeland degradation, the effects on the cumulative loss in agricultural Gross Domestic Product (GDP) and on overall poverty at a national level can therefore be enormous. For example, in South Africa, the success of industrial development depends partially on the agricultural sector and its improvements (Poonyth et al. 2001), and about 10% of formal employment is related to agriculture (RSA 2017). Also in Namibia, agriculture is a key sector of the economy. It is not only the largest employer but also critical to livelihoods and food security (MET 2020).
7 Strategies to Mitigate the Effects of Degradation
One of the most effective ways to reduce pressure on degraded rangelands is to reduce livestock numbers by selling animals. However, destocking has not been a common strategy to cope with limited grazing resources on communal land, mainly due to social prestige and wealth attached to having a large herd of cattle. Traditionally, one of the main strategies to deal with drought and declining grazing resources in communal areas has been to move livestock (particularly cattle) from degraded to nondegraded areas during the dry season (Twyman et al. 2001). This practice is still being largely done especially in the north-eastern parts of Namibia and involves herders moving herds of cattle over long distances in search for fodder. Since the twentieth century, many parts of Namibia have seen restricted movement of cattle, mainly due to limited land and policies leading to overpopulation and restricted red meat transfer between regions (see Sect. 15.4.2). After independence, government subsidies have also enabled people to keep livestock without the necessity to move them. This encourages many livestock farmers to keep animals during periods of fodder scarcity, resulting in further degradation of rangelands.
Yet, there have been some advances in successful mitigation of rangeland degradation: To detain bush encroachment and promote de-bushing, a growing industry of bush utilization can benefit farmers using it for economic gain; either as possible animal fodder in times with scarce resources or for charcoal production. The goal is mostly reducing rather than eliminating woody species completely (de Klerk 2004; Haussmann et al. 2016). With sustainable bush harvesting, the former threat of bush encroachment has become an important opportunity for economic income. In Namibia, more than 5000 jobs have been created in the biomass sector initiated by development projects (e.g., Bush Control and Biomass Utilization, BCBU) since 2015 (GIZ 11/10/2020). This has also led to an increase in wages and created a new opportunity for rural employment. With at least 2.6% of global charcoal export (USD 34.1 million in 2018), Namibia has reached the top rank in Africa and rank 12 word-wide (South Africa with 1.5%: 2nd in Africa, 18th world-wide) (Shikangalah and Mapani 2020). It is also estimated that in Namibia, the plain utilization of firewood (e.g., from bushes) currently ranges between 0.5 and 1 million tons per year and is projected to reach 1.2 million in 2025 (Stafford et al. 2017). The future use of these large-scale de-bushed areas, however, remains uncertain. An immediate return to livestock production is difficult and requires well planned restoration and aftercare measures, since viable grass seeds in the soil are lacking, the reintroduction of native savanna grasses is extremely difficult (van den Berg and Kellner 2005; Kinyua et al. 2010), and bushes often resprout (Bhattachan et al. 2014). Resaturation includes active and passive options such as furrowing, brush packing and revegetation (e.g., van den Berg and Kellner 2005; Harmse et al. 2016). Bush control techniques by prescribed fire, as well as manual and chemical methods (e.g., Trollope 2011; Joubert et al. 2012; Lohmann et al. 2014) are aftercare measurements to prevent resprouting and reestablishment of bushes. However, the potential impacts of these measures on biodiversity and ecosystem functioning remain topics of active debate and research.
Wildlife-based management as a strategy to approach rangeland degradation is increasingly applied but not well understood. Increased browsing of wildlife on seedlings and saplings has the potential to mitigate bush encroachment and to contribute toward conservation (McGranahan 2008; Irob et al. 2022). Additionally, wildlife has shown more resilience to local environmental change compared to livestock, which might make it a better management strategy in the face of degradation. The wide forage selection of wild herbivores allows them to cope with scarcity in palatable grass species in degraded lands (Taylor and Walker 1978, cited in McGranahan 2008). Growing evidence suggests that wildlife-based land uses offer higher socioeconomic benefits compared to livestock in semiarid rangelands in both Namibia and South Africa (Lindsey et al. 2013). There is consensus on the potential economic benefits that wildlife can provide through consumptive and nonconsumptive utilization (Lindsey et al. 2013; NACSO 2019).
As a response to changing environmental and socioeconomic conditions, a shift from cattle farming to wildlife farming has been observed in South African rangelands since the 1960 (Chaminuka et al. 2012) and more recently in Namibia. While this shift in land use can be profitable on commercial freehold farms, such a shift can be challenging in other communities and communal managed systems. Since wildlife is a common pool resource in communal areas, the allocation of direct benefits (e.g., income) is much harder to implement. In Namibia, the presence of Community Based Natural Resource Management (CBNRM) such as conservancies and community forests covering about 22% of the total land area (NACSO 2019) provides however an opportunity to develop wildlife management as a land use option to deal with rangeland degradation. Whereas Namibia has community-based conservation areas, where communities can manage and benefit from wildlife, more reforms in land tenure rights may be required in countries where such legal frameworks are still in progress.
Evidence from ecological studies suggests that individuals and communities can successfully reduce land degradation (Herrick et al. 2013), and restoration might be possible. However, mitigation measures can be very expensive, elaborate and local people might often not have the capacity to implement them sufficiently without support (Bourne et al. 2017). This is either because financial support is lacking (Inman 2020) or because the knowledge about these measures or how to implement them correctly is missing. Additionally, evidence suggests that complete recovery will hardly be achieved (Parkhurst et al. 2021) especially in systems where overgrazing is a continuous threat. Proper, institutionalized aid might promote restoration and could potentially advance resilience (Bourne et al. 2017).
8 Conclusions
-
1.
Rangeland degradation is an ongoing threat to many savannas in Southern Africa, which is not only expressed through loss of fodder and soil. Symptoms comprise bush encroachment, lowered groundwater tables, reduced water quality, soil fertility, invasive species and both loss of—and changes in—plant and animal diversity. Neither ecological nor socioeconomic knowledge alone is sufficient to understand complex socioecological relationships and effectively manage savanna rangeland systems.
-
2.
Concerns and actions related to the loss of forage production should be broadened to include biodiversity and its multiple functional relationships with the water and nutrient cycle, primary production and trophic interactions.
-
3.
It is important to strengthen alternative strategies for conserving and restoring ecosystems and improving human well-being, for example, by implementing local solutions through community-based grazing management (e.g., communal conservancies) and land use diversification.
-
4.
A shift from cattle farming to wildlife farming has the potential to contribute toward biodiversity conservation, the control of species causing bush-encroachment and increased economic benefits for local communities. We note, however, that more scientific evidence, reforms in land tenure rights and improved restoration practices are needed.
References
ABSA (2003) Game ranch profitability in Southern Africa. Rivonia, the SA. Financial Sector Forum: 73 pp
Allen VG et al (2011) An international terminology for grazing lands and grazing animals. Grass Forage Sci 66(1):2–28. https://doi.org/10.1111/j.1365-2494.2010.00780.x
Angassa A (2005) The ecological impact of bush encroachment on the yield of grasses in Borana rangeland ecosystem. Afr J Ecol 43:14–20. https://doi.org/10.1111/j.1365-2028.2005.00429.x
Archer S, Schimel DS, Holland EA (1995) Mechanisms of shrub land expansion: land use, climate or CO2. Clim Chang 29:91–99
Archer SR, Andersen EM, Predick KI, Schwinning S, Steidl RJ, Woods SR (2017) Woody plant encroachment: causes and consequences. In: Briske DD (ed) Rangeland systems. Springer International Publishing, pp 25–84. https://doi.org/10.1007/978-3-319-46709-2_2
Banerjee AV, Duflo E (2007) The economic lives of the poor. J Econ Perspect 21(1):141–167. https://doi.org/10.1257/jep.21.1.141
Barbier EB, Hochard JP (2018) Land degradation and poverty. Nat Sustain 1(11):623–631. https://doi.org/10.1038/s41893-018-0155-4
Bargués Tobella A, Reese H, Almaw A, Bayala J, Malmer A, Laudon H, Ilstedt U (2014) The effect of trees on preferential flow and soil infiltrability in an agroforestry parkland in semiarid Burkina Faso. Water Resour Res 50(4):3342–3354. https://doi.org/10.1002/2013WR015197
Barrett CB, Bevis LEM (2015) The self-reinforcing feedback between low soil fertility and chronic poverty. Nat Geosci 8(12):907–912. https://doi.org/10.1038/ngeo2591
Baumert S et al (2016) Charcoal supply chains from Mabalane to Maputo: who benefits? Energy Sustain Dev 33:129–138. https://doi.org/10.1016/j.esd.2016.06.003
Beinart W (2003) The rise of conservation in South Africa. Settlers, livestock, and the environment 1770–1950. Oxford University Press, Oxford. checked on 8/3/2021
Belay TA, Totland Ø, Moe SR (2013) Ecosystem responses to woody plant encroachment in a semiarid savanna rangeland. Plant Ecol 214:1211–1222. https://doi.org/10.1007/s11258-013-0245-3
Belsky AJ (1994) Influences of trees on savanna productivity: tests of shade, nutrients, and treegrass competition. Ecology 75:922–932
Berzborn S, Solich M (2013) Pastoralism and nature conservation in Southern Africa. In: Bollig M, Schnegg M, Wotzka H-P (eds) Pastoralism in Africa. Past, present, and future. Berghahn, New York, pp 440–472
Beyer M, Hamutoko JT, Wanke H, Gaj M, Koeniger P (2018) Examination of deep root water uptake using anomalies of soil water stable isotopes, depth-controlled isotopic labeling and mixing models. J Hydrol 566:122–136
Bhattachan A, D’Odorico P, Dintwe K, Okin GS, Collins SL (2014) Resilience and recovery potential of duneland vegetation in the southern Kalahari. Ecosphere 5:2–14
Blaum N, Rossmanith E, Popp A, Jeltsch F (2007a) Shrub encroachment affects mammalian carnivore abundance and species richness in semiarid rangelands. Acta Oecol 31:86–92
Blaum N, Rossmanith E, Schwager M, Jeltsch F (2007b) Responses of mammalian carnivores to land use in arid Kalahari rangelands. Basic Appl Ecol 8:552–564
Blaum N, Rossmanith E, Jeltsch F (2007c) Land use affects rodent communities in Kalahari savannah rangelands. Afr J Ecol 45:189–195
Blaum N, Seymour C, Rossmanith E, Schwager M, Jeltsch F (2009) Changes in arthropod diversity along a land use driven gradient of shrub cover in savanna rangelands: identification of suitable indicators. Biodivers Conserv 18(5):1187–1199. https://doi.org/10.1007/s10531-008-9498-x
Blumröder J, Eccard J, Blaum N (2012) Behavioural flexibility in foraging mode of the spotted sand lizard (Pedioplanis l. lineoocellata) seems to buffer negative impacts of savanna degradation. J Arid Environ 77:149–152
Bollig M (2013) Chapter 10: Social-ecological change and institutional development in a pastoral Community in North-western Namibia. In: Bollig M, Schnegg M, Wotzka H-P (eds) Pastoralism in Africa. Past, present, and future. Berghahn, New York, pp 316–340
Bollig M, Osterle M (2008) Changing communal land tenure in an East African pastoral system: institutions and socio-economic transformations among the Pokot of NW Kenya. Z Ethnol 133(2):301–322
Bonachela JA, Pringle RM, Sheffer E, Coverdale TC, Guyton JA, Caylor KK, Levin SA, Tarnita CE (2015) Termite mounds can increase the robustness of dryland ecosystems to climatic change. Science 347(6222):651–655. https://doi.org/10.1126/science.1261487
Bond I (2001) CAMPFIRE and the incentives for institutional change, in African wildlife and livlihoods. The promise and performance of community conservation. James Currey, Oxford
Bond WJ, Midgley GF (2012) Carbon dioxide and the uneasy interaction of trees and savannah grasses. Philos Trans R Soc Lond B Biol Sci 367:601–612
Bourne A, Muller H, De Villiers A, Alam M, Hole D (2017) Assessing the efficiency and effectiveness of rangeland restoration in Namaqualand, South Africa. Plant Ecol 218(1):7–22. https://doi.org/10.1007/s11258-016-0644-3
Breytenbach W (2004) Land reform in Southern Africa. In: Hunter J (ed) Who should own the land? Analyses and views on land reform and the land question in Namibia and southern Africa. Konrad-Adenauer-Stiftung; Namibia Institute for Democracy, Windhoek Namibia, pp 46–63
Brown J, Scholtz CH, Janeau J-L, Grellier S, Podwojewski P (2010) Dung beetles (Coleoptera: Scarabaeidae) can improve soil hydrological properties. Appl Soil Ecol 46(1):9–16. https://doi.org/10.1016/j.apsoil.2010.05.010
Buisson E, Stradic SL, Silveira FAO, Durigan G, Overbeck GE, Fidelis A, Fernandes GW, Bond WJ, Hermann J-M, Mahy G, Alvarado ST, Zaloumis NP, Veldman JW (2019) Resilience and restoration of tropical and subtropical grasslands, savannas, and grassy woodlands. Biol Rev 94(2):590–609. https://doi.org/10.1111/brv.12470
Cammeraat LH, Willott SJ, Compton SG, Incoll LD (2002) The effects of ants’ nests on the physical, chemical and hydrological properties of a rangeland soil in semi-arid Spain. Geoderma 105(1):1–20. https://doi.org/10.1016/S0016-7061(01)00085-4
Carruthers J (2008) “Wilding the farm or farming the wild”? The evolution of scientific game ranching in South Africa from the 1960s to the present. Trans R Soc South Africa 63(2):160–181. https://doi.org/10.1080/00359190809519220
Chaminuka P, McCrindle CME, Udo HMJ (2012) Cattle farming at the wildlife/livestock Interface: assessment of costs and benefits adjacent to Kruger National Park, South Africa. Soc Nat Resour 25(3):235–250. https://doi.org/10.1080/08941920.2011.580417
Chown SL (2010) Temporal biodiversity change in transformed landscapes: a southern African perspective. Philos Trans R Soc Lond B Biol Sci 365:3729–3742
Colloff MJ, Pullen KR, Cunningham SA (2010) Restoration of an ecosystem function to revegetation communities: the role of invertebrate macropores in enhancing soil water infiltration. Restor Ecol 18:65–72. https://doi.org/10.1111/j.1526-100X.2010.00667.x
Dahlberg AC (2000) Landscape(s) in transition: an environmental history of a village in north-East Botswana. J South Afr Stud 26(4):759–782. https://doi.org/10.1080/03057070020008260
de Klerk JN (2004) Bush encroachment in Namibia. Report on phase 1 of the bush encroachment research, monitoring, and management project. Edited by Ministry of Environment and Tourism, Government of the Republic of Namibia. Published. John Meinert Printers Windhoek
Delius P, Schirmer S (2000) Soil conservation in a racially ordered society: South Africa 1930–1970. J South Afr Stud 26(4):719–742. https://doi.org/10.1080/713683610
Demas D, Sylvia D, Nancy M, Forman S, Jaap A, Stephane F, Philip S (2012) Livestock competitiveness, economic growth and opportunities for job creation in Namibia
Diao X, Hazell P, Thurlow J (2010) The role of agriculture in African development. World Dev 38(10):1375–1383. https://doi.org/10.1016/j.worlddev.2009.06.011
Dlamini P, Mbanjwa V, Gxasheka M, Tyasi L, Sekhohola-Dlamini L (2019) Chemical stabilisation of carbon stocks by polyvalent cations in plinthic soil of a shrub-encroached savanna grassland, South Africa. Catena 181:104088. https://doi.org/10.1016/j.catena.2019.104088
Dovie DBK, Witkowski ETF, Shackleton CM (2004) The fuelwood crisis in Southern Africa – relating fuelwood use to livelihoods in a Rural Village. GeoJournal 60:123–133. https://doi.org/10.1023/B:GEJO.0000033597.34013.9f
Dreber N, van Rooyen SE, Kellner K (2018) Relationship of plant diversity and bush cover in rangelands of a semi-arid Kalahari savannah, South Africa. Afr J Ecol 56:132–135. https://doi.org/10.1111/aje.12425
Eldridge DJ, Bowker MA, Maestre FT, Roger E et al (2011) Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecol Lett 14:709–722
Ellery WN, Scholes RJ, Mentis MT (1991) An initial approach to predicting the sensitivity of the South African grassland biome to climate change. S Afr J Sci 87:499–503
Ellis EC, Ramankutty N (2008) Putting people in the map: anthropogenic biomes of the world. Front Ecol Environ 6:439–447. https://doi.org/10.1890/070062
Funk C, Peterson P, Landsfeld M, Pedreros D, Verdin J, Shukla S et al (2015) The climate hazards infrared precipitation with stations—a new environmental record for monitoring extremes. Sci Data 2(1):1–21
Geissler K, Hahn C, Joubert D, Blaum N (2019a) Functional responses of the herbaceous plant community explain ecohydrological feedbacks of savanna shrub encroachment. Perspect Plant Ecol Evol Syst 39:125458. https://doi.org/10.1016/j.ppees.2019.125458
Geissler K, Heblack J, Uugulu S, Wanke H, Blaum N (2019b) Partitioning of water between differently sized shrubs and potential groundwater recharge in a semiarid savanna in Namibia. Front Plant Sci 10. https://doi.org/10.3389/fpls.2019.01411
GIZ (11/10/2020): Namibia: Bush waste boosts the economy. Laufs, Johannes. Deutsche Gesellschaft für Internationale Zusammenarbeit. Available online at https://www.giz.de/en/workingwithgiz/91230.html, checked on 7/26/2021
Gomiero T (2016) Soil degradation, land scarcity and food security: reviewing a complex challenge. Sustainability 8(3):281. https://doi.org/10.3390/su8030281
Groengroeft A, de Blécourt M, Classen N, Landschreiber L, Eschenbach A (2018) Acacia trees modify soil water dynamics and the potential groundwater recharge in savanna ecosystems. In: Revermann R, Krewenka KM, Schmiedel U, Olwoch JM, Helmschrot J, Jürgens N (eds) Climate change and adaptive land management in southern Africa – assessments, changes, challenges, and solutions, Biodiversity & Ecology, vol 6. Klaus Hess Publishers, Göttingen & Windhoek, pp 177–186. https://doi.org/10.7809/b-e.00321
Harmse CJ, Kellner K, Dreber N (2016) Restoring productive rangelands: a comparative assessment of selective and non-selective chemical bush control in a semi-arid Kalahari savanna. J Arid Environ 135:39–49
Hattingh ER (1952) Comments on thorn scrub control with herbicides in Africa. Weeds 1(4):372–373
Haussmann NS, Kalwij JM, Bezuidenhout S (2016) Some ecological side-effects of chemical and physical bush clearing in a southern African rangeland ecosystem. S Afr J Bot 102:234–239. https://doi.org/10.1016/j.sajb.2015.07.012
Hering R, Hauptfleisch M, Geißler K, Marquart A, Schoenen M, Blaum N (2019) Shrub encroachment is not always land degradation: insights from ground-dwelling beetle species niches along a shrub cover gradient in a semi-arid Namibian savanna. Land Degrad Dev 30(1):14–24. https://doi.org/10.1002/ldr.3197
Herrick JE, Sala OE, Karl JW (2013) Land degradation and climate change: a sin of omission? Front Ecol Environ 11(6):283. https://doi.org/10.1890/1540-9295-11.6.283
Higgins SI, Bond WJ, Trollope WSW (2000) Fire, resprouting and variability: a recipe for grass-tree coexistence in savanna. J Ecol 88(2):213–229. https://doi.org/10.1046/j.1365-2745.2000.00435.x
Hiscock KM, Lloyd JW, Lerner DN (1991) Review of natural and artificial denitrification of groundwater. Water Res 25(9):1099–1111
Hoffman MT (2014) Changing patterns of rural land use and land cover in South Africa and their implications for land reform. J South Afr Stud 40(4):707–725. https://doi.org/10.1080/03057070.2014.943525
Hoffman MT, Todd S (2000) A national review of land degradation in South Africa: the influence of biophysical and socio-economic factors. J South Afr Stud 26(4):743–758. https://doi.org/10.1080/713683611
Honsbein D, Shiningavamwe K, Iikela J, de la Puerta Fernandez ML (2017) Animal feed from Namibian encroacher bush. Available online at https://www.dasnamibia.org/download/brochures/GIZ-UNDP-MAWF-Animal-Feed-Manual-2017.pdf
Humphrey GL, Gillson L, Ziervogel G (2021) How changing fire management policies affect fire seasonality and livelihoods. Ambio 50:475–491
Huntley BJ (1982) Southern African savannas. In: Huntley BJ, Walker BH (eds) Ecology of tropical savannas. Springer-Verlag, Berlin Heidelberg. ISBN 978-3-642-68786-0
Inman EN (2020) Community conservation and restoration of degraded land in semi-arid Namibia in the context of climate change. Dissertation, University of Western Australia. School of Biological Sciences, checked on 8/3/2021
Irob K, Blaum N, Baldauf S, Kerger L, Strohbach B, Kanduvarisa A, Lohmann D, Tietjen B (2022). Browsing herbivores improve the state and functioning of savannas: a model assessment of alternative land use strategies. https://doi.org/10.22541/au.162682707.72443437/v1
Joubert DF, Rothauge A, Smit GN (2008) A conceptual model of vegetation dynamics in the semiarid Highland savanna of Namibia, with particular reference to bush thickening by Acacia mellifera. J Arid Environ 72:2201–2210. https://doi.org/10.1016/j.jaridenv.2008.07.004
Joubert DF, Smit GN, Hoffman MT (2012) The role of fire in preventing transitions from a grass dominated state to a bushthickened state in arid savannas. J Arid Environ 87:1e7
Kaiser D, Lepage M, Konaté S, Linsenmair KE (2017) Ecosystem services of termites (Blattoidea: Termitoidae) in the traditional soil restoration and cropping system Zaï in Northern Burkina Faso (West Africa). Agric Ecosyst Environ 236(Supplement C):198–211. https://doi.org/10.1016/j.agee.2016.11.023
Kashululu R-MP, Paul H (2020) Chapter 8: The fencing question in Namibia: a case study in Omusati region. In: Odendaal W, Werner W (eds) “Neither here nor there”. Indigeneity, marginalisation and land rights in post-independence Namibia. Legal Assistance Centre, Windhoek Namibia, pp 163–182. Available online at https://library.wur.nl/webquery/wurpubs/fulltext/521219
Kinyua D, McGeoch LE, Georgiadis N, Young TP (2010) Short-term and long-term effects of soil ripping, seeding and fertilization on the restoration of a tropical rangeland. Restor Ecol 18:226–233
Kreike E (2009) De-globalisation and deforestation in colonial Africa: closed markets, the cattle complex, and environmental change in North-Central Namibia, 1890–1990. J South Afr Stud 35(1):81–98. https://doi.org/10.1080/03057070802685585
Kulmatiski A, Beard KH, Verweij RJT, February EC (2010) A depth-controlled tracer technique measures vertical, horizontal and temporal patterns of water use by trees and grasses in a subtropical savanna. New Phytol 188:199–209. https://doi.org/10.1111/j.1469-8137.2010.03338.x
Lal R (2015) Restoring soil quality to mitigate soil degradation. Sustainability 7(5):5875–5895. https://doi.org/10.3390/su7055875
Lange G-M, Barnes JI, Motinga DJ (1998) Cattle numbers, biomass, productivity and land degradation in the commercial farming sector of Namibia, 1915-95. Dev South Afr 15(4):555–572. https://doi.org/10.1080/03768359808440031
Lee E, Kumar P, Barron G, Greg A, Hendryx SM (2018) Impact of hydraulic redistribution on multispecies vegetation water use in a semiarid savanna ecosystem: an experimental and modeling synthesis. Water Resour Res 54(6):4009–4027. https://doi.org/10.1029/2017WR021006
Léonard J, Perrier E, Rajot JL (2004) Biological macropores effect on runoff and infiltration: a combined experimental and modelling approach. Agric Ecosyst Environ 104(2):277–285. https://doi.org/10.1016/j.agee.2003.11.015
Li H, Shen H, Chen L, Liu T, Hu H, Zhao X, Zhou L, Zhang P, Fang J (2016) Effects of shrub encroachment on soil organic carbon in global grasslands. Sci Rep 6(1):28974. https://doi.org/10.1038/srep28974
Lindsey PA, Havemann CP, Lines RM, Price AE, Retief TA, Rhebergen T, Van der Waal C, Romañach SS (2013) Benefits of wildlife-based land uses on private lands in Namibia and limitations affecting their development. Oryx 47(01):41–53. https://doi.org/10.1017/S0030605311001049
Lohmann D, Tietjen B, Blaum N, Joubert DF, Jeltsch F (2014) Prescribed fire as a tool for managing shrub encroachment in semi-arid savanna rangelands. J Arid Environ 107:49–56. https://doi.org/10.1016/j.jaridenv.2014.04.003
Luoga EJ, Witkowski ETF, Balkwill K (2000) Economics of charcoal production in miombo woodlands of eastern Tanzania: some hidden costs associated with commercialization of the resources. Ecol Econ 35(2):243–257. https://doi.org/10.1016/S0921-8009(00)00196-8
Mander M (1998) Marketing of indigenous medicinal plants in South Africa: a case study in Kwazulu-Natal. Food and Agriculture Organization of the United Nations, Rome. https://doi.org/10.13140/2.1.1073.4084
Marchiori L, Maystadt J-F, Schumacher I (2012) The impact of weather anomalies on migration in sub-Saharan Africa. J Environ Econ Manage 63(3):355–374. https://doi.org/10.1016/j.jeem.2012.02.001
Marquart A, Eldridge DJ, Geissler K, Lobas C, Blaum N (2020a) Interconnected effects of shrubs, invertebrate-derived macropores and soil texture on water infiltration in a semi-arid savanna rangeland. Land Degrad Dev 31(16):2307–2318. https://doi.org/10.1002/ldr.3598
Marquart A, Geissler K, Heblack J, Lobas C, Münch E, Blaum N (2020b) Individual shrubs, large scale grass cover and seasonal rainfall explain invertebrate-derived macropore density in a semi-arid Namibian savanna. J Arid Environ 176:104101. https://doi.org/10.1016/j.jaridenv.2020.104101
Marquart A, Goldbach L, Blaum N (2020c) Soil-texture affects the influence of termite macropores on soil water infiltration in a semi-arid savanna. Ecohydrology 13(8):e2249. https://doi.org/10.1002/eco.2249
McGranahan DA (2008) Managing private, commercial rangelands for agricultural production and wildlife diversity in Namibia and Zambia. Biodivers Conserv 17(8):1965–1977. https://doi.org/10.1007/s10531-008-9339-y
Mcleon G (1995) Environmental change at Letlhakeng in the Kweneng District. Botsw Notes Rec 27:81–298
McNaughton SJ, Ruess RW, Seagle SW (1988) Large mammals and process dynamics in African ecosystems: herbivorous mammals affect primary productivity and regulate recycling balances. Bioscience 38:794–800. https://doi.org/10.2307/1310789
Meik JM, Jeo RM, Mendelson JR, Jenks KE (2002) Effects of bush encroachment on an assemblage of diurnal lizard species in central Namibia. Biol Conserv 106:29–36
Menestrey Schwieger DA, Mbidzo M (2020) Socio-historical and structural factors linked to land degradation and desertification in Namibia’s former Herero ‘homelands’. J Arid Environ 178:1–7. https://doi.org/10.1016/j.jaridenv.2020.104151
MET (2020) Fourth National Communication to the United Nations Framework Convention on Climate Change. Edited by Ministry of Environment and Tourism. Government of the Republic of Namibia, Windhoek
Metzger JC, Landschreiber L, Grongroft A, Eschenbach A (2014) Soil evaporation under different types of land use in southern African savanna ecosystems. J Plant Nutr Soil Sci 177:468–475. https://doi.org/10.1002/jpln.201300257
Mogashoa R, Dlamini P, Gxasheka M (2021) Grass species richness decreases along a woody plant encroachment gradient in a semi-arid savanna grassland, South Africa. Landsc Ecol 36:617–636. https://doi.org/10.1007/s10980-020-01150-1
Moore A, Odendaal A (1987) The economic implications of bush encroachment and bush control in a weaner calf production system in the Thorny Bushveld of the Molopo area. Afr J Range Forage Sci 4(4)
Mucina L, Rutherford MC (eds) (2006) The vegetation of South Africa, Lesotho and Swaziland. South African National Biodiversity Institute (Strelitzia), Pretoria, p 19
Mudombi S et al (2018) User perceptions about the adoption and use of ethanol fuel and cookstoves in Maputo, Mozambique. Energy Sustain Dev 44:97–108. https://doi.org/10.1016/j.esd.2018.03.004
Mukwada G (2018) Savanna ecosystems of southern Africa. In: Holmes PJ, Boardman J (eds) Southern landscapes and environmental change. Taylor and Francis. https://doi.org/10.4324/9781315537979-10
Muntifering JR, Dickman AJ, Perlow LM et al (2006) Managing the matrix for large carnivores: a novel approach and perspective from cheetah (Acinonyx jubatus) habitat suitability modelling. Anim Conserv 9:103–112
NACSO (2019) State of community conservation in Namibia: a review of communal conservancies, community forests and other CBNRM initiatives (p. 88) [Annual report]. http://www.nacso.org.na/resources/state-of-community-conservation
NAU (Namibia Agricultural Union) (2010) The effect of bush encroachment on groundwater resources in Namibia: a desk top study (Issue December)
Nichols E, Spector S, Louzada J, Larsen T, Amezquita S, Favila ME (2008) Ecological functions and ecosystem services provided by Scarabaeinae dung beetles. Biol Conserv 141(6):1461–1474. https://doi.org/10.1016/j.biocon.2008.04.011
Nkem JN, Lobry de Bruyn LA, Grant CD, Hulugalle NR (2000) The impact of ant bioturbation and foraging activities on surrounding soil properties. Pedobiologia 44(5):609–621. https://doi.org/10.1078/S0031-4056(04)70075-X
O'Connor TG, Puttick JR, Timm Hoffman TM (2014) Bush encroachment in southern Africa: changes and causes. Afr J Range Forage Sci 31(2):67–88. https://doi.org/10.2989/10220119.2014.939996
Olson DM et al (2001) Terrestrial ecoregions of the world: a new map of life on earth. Bioscience 51(11):933. https://doi.org/10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2
Parkhurst T, Prober SM, Hobbs RJ, Standish RJ (2021) Global meta-analysis reveals incomplete recovery of soil conditions and invertebrate assemblages after ecological restoration in agricultural landscapes. J Appl Ecol. https://doi.org/10.1111/1365-2664.13852
Poonyth D, Hassan R, Kirsten JF, Calcaterra M (2001) Is agricultural sector growth a precondition for economic growth? The case of South Africa. Agrekon 40(2):269–279. https://doi.org/10.1080/03031853.2001.9524950
Potts D (2012) Land alienation under colonial and white settler governments in southern Africa historical land ‘grabbing’. In: Allan T, Keulertz M, Sojamo S, Warner J (eds) Handbook of land and water grabs in Africa. Foreign direct investment and food and water security, 1st edn. Routledge, Abingdon, pp 24–42. checked on 8/3/2021
Reed MS, Stringer LC, Dougill AJ, Perkins JS, Atlhopheng JR, Mulale K, Favretto N (2015) Reorienting land degradation towards sustainable land management: linking sustainable livelihoods with ecosystem services in rangeland systems. J Environ Manag 151:472–485. https://doi.org/10.1016/j.jenvman.2014.11.010
Rietkerk M, van de Koppel J (1997) Alternate stable states and threshold effects in semi-arid grazing systems. Oikos 79:69–76
Rohde RF, Hoffman MT (2012) The historical ecology of Namibian rangelands: vegetation change since 1876 in response to local and global drivers. Sci Total Environ 416:276–288. https://doi.org/10.1016/j.scitotenv.2011.10.067
RSA (ed) (2017) Agriculture and land. National Treasury, Republic of South Africa, Pretoria. Chapter 9; (Provincial Budgets and Expenditure Review 2010/11–2016/17)
Rückamp D, Martius C, Bornemann L, Kurzatkowski D, Naval LP, Amelung W (2012) Soil genesis and heterogeneity of phosphorus forms and carbon below mounds inhabited by primary and secondary termites. Geoderma 170:239–250. https://doi.org/10.1016/j.geoderma.2011.10.004
Sanchez P, Palm C, Sachs J, Denning G, Flor R, Harawa R et al (2007) The African millennium villages. Proc Natl Acad Sci USA 104(43):16775–16780. https://doi.org/10.1073/pnas.0700423104
Sandhage-Hofmann A, Löffler J, Kotze E, Weijers S, Wingate V, Wundram D, Weihermüller L, Pape R, du Preez CC, Amelung W (2020) Woody encroachment and related soil properties in different tenure-based management systems of semiarid rangelands. Geoderma 372:114399
Sankaran M, Hanan NP, Scholes RJ, Ratnam J, Augustine DJ, Cade BS, Gignoux J et al (2005) Determinants of Woody Cover in African Savannas. Nature 438. (December 2005):846–849. https://doi.org/10.1038/nature04070
Schnegg M, Pauli J, Greiner C (2013) Chapter 11: Pastoral belonging: causes and consequences of part-time pastoralism in North-Western Namibia. In: Bollig M, Schnegg M, Wotzka H-P (eds) Pastoralism in Africa. Past, present, and future. Berghahn, New York, pp 341–364
Schneider HP (2012) The history of veterinary medicine in Namibia. J S Afr Vet Assoc 83(1):1–11. https://doi.org/10.4102/jsava.v83i1.4
Scholes RJ (2004) Savanna. In: Cowling RM, Richardson DM, Pierce SM (eds) Vegitation of Southern Africa. Cambridge University Press, Cambridge
Scholes RJ, Archer SR (1997) Tree-grass interactions in savannas. Annu Rev Ecol Syst 28:545–570
Shackleton CM (1993) Are the communal grazing lands in need of saving? Dev South Afr 10(1):65–78. https://doi.org/10.1080/03768359308439667
Shackleton C, Shackleton S (2004) The importance of non-timber forest products in rural livelihood security and as safety nets: a review of evidence from South Africa. S Afr J Sci 100:558–664
Shackleton C, Shackleton S, Gambiza J, Nel E, Rowntree K, Urquhart P (2008) Links between ecosystem services and poverty alleviation. Situation analysis for arid and semi-arid lands in southern Africa. Edited by Consortium on Ecosystems and Poverty in Sub-Saharan Africa (CEPSA)
Shackleton S, Cocks M, Dold T, Kaschula S, Mbata K, Mickels-Kokwe G, von Maltitz GP (2010) Non-wood forest products: description, use and management. In: Chidumayo EN, Gumbo DJ (eds) The dry forests and woodlands of Africa: managing for products and services. Earthscan, London
Shikangalah RN, Mapani BS (2020) A review of bush encroachment in Namibia: from a problem to an opportunity? J Rangel Sci 10(3):251–266
Sirami C, Seymour C, Midgley G, Barnard P (2009) The impact of shrub encroachment on savanna bird diversity from local to regional scale. Divers Distrib 15:948–957
Sitters J, Edwards PJ, Venterink HO (2013) Increases of soil C, N, and P pools along an acacia tree density gradient and their effects on trees and grasses. Ecosystems 16:347–357
Smit GN (2004) An approach to tree thinning to structure southern African savannas’ for long-term restoration from bush encroachment. J Environ Manag 71:179–191. https://doi.org/10.1016/j.jenvman.2004.02.005
Smit IPJ, Prins HHT (2015) Predicting the effects of woody encroachment on mammal communities, grazing biomass and fire frequency in African savannas. PLoS One 10:e0137857
Stafford W, Birch C, Etter H, Blanchard R, Mudavanhu S, Angelstam P, Blignaut J, Ferreira L, Marais C (2017) The economics of landscape restoration: benefits of controlling bush encroachment and invasive plant species in South Africa and Namibia. Ecosyst Serv 27:1–10. https://doi.org/10.1016/j.ecoser.2016.11.021
Stevens N, Erasmus BFN, Archibald S, Bond WJ (2016) Woody encroachment over 70 years in South African savannahs: overgrazing, global change or extinction aftershock? Philos Trans R Soc B 371(1703):20150437. https://doi.org/10.1098/rstb.2015.0437
Suich H, Howe C, Mace G (2015) Ecosystem services and poverty alleviation: a review of the empirical links. Ecosyst Serv 12:137–147. https://doi.org/10.1016/j.ecoser.2015.02.005
Tallis H, Kareiva P (2007) Ecosystem services. Curr Biol 15(18):R746–R748
Traore S, Tigabu M, Jouquet P, Ouedraogo SJ, Guinko S, Lepage M (2015) Long-term effects of Macroternies termites, herbivores and annual early fire on woody undergrowth community in Sudanian woodland, Burkina Faso. Flora 211:40–50. https://doi.org/10.1016/j.flora.2014.12.004
Trede R, Patt R (2015) Value added end-use opportunities for Namibian encroacher bush
Trollope WSW, Dondofema F (2003) Role of fire, continuous browsing and grazing in controlling bush encroachment in the arid savannas of the Eastern Cape province in South Africa. In: Allsopp N, Palmer AR, Milton SJ, Kerley GIH, Kirkman KP, Hurt R, Brown C (eds) Rangelands in the new millennium: Proceedings of the 7th international rangeland conference, Durban, South Africa. International Rangeland Congress, Durban, pp 408–411
Tully K, Sullivan C, Weil R, Sanchez P (2015) The state of soil degradation in sub-Saharan Africa: baselines, trajectories, and solutions. Sustainability 7(6):6523–6552. https://doi.org/10.3390/su7066523
Twyman C, Dougill A, Sporton D, Thomas D (2001) Community fencing in open rangelands: self-empowerment in Eastern Namibia. Rev Afr Polit Econ 28(87):9–26. https://doi.org/10.1080/03056240108704500
Uugulu, S (2022) Estimation of groundwater recharge along a precipitation gradient for Savanna aquifers in Namibia. PhD Thesis, University of Namibia, 180 pages
van den Berg L. and Kellner K. (2005) Restoring degraded patches in a semi-arid rangeland of South Africa. J. Arid Environ 6::497–511. https://doi.org/10.1016/j.jaridenv.2004.09.024
Vehrs HP, Heller GR (2017) Fauna, fire, and farming: landscape formation over the past 200 years in pastoral East Pokot, Kenya. In: Gale Group (ed) Human ecology. Springer New York LLC, New York. https://doi.org/10.1007/s10745-017-9926-1
Walter H (1939) Grassland, Savanne und Busch der arideren Teile Afrikas in ihrer ökologischen Bedingtheit. Jahrbücher für wissenschaftliche Botanik 87:750–860
Ward D (2005) Do we understand the causes of bush encroachment in African savannas? Afr J Range Forage Sci 22(2):101–105
Wasiolka B, Blaum N (2011) Comparing biodiversity between protected savanna and adjacent non-protected farmland in the southern Kalahari. J Arid Environ 75:836–841
Weitzman JN, Brooks JR, Mayer PM, Rugh WD, Compton JE (2021) Coupling the dual isotopes of water (δ 2 H and δ 18 O) and nitrate (δ 15 N and δ 18 O): a new framework for classifying current and legacy groundwater pollution. Environ Res Lett 16(4):1
Wigley BJ, Bond WJ, Hoffman MT (2009) Bush encroachment under three contrasting land-use practices in a Mesic South African savanna. Afr J Ecol 47(SUPPL. 1):62–70. https://doi.org/10.1111/j.1365-2028.2008.01051.x
Yeaton RI (1988) Porcupines, fires and the dynamics of the tree layer of Burkea africana savanna. J Ecol 76:1017–1029
Yuan C, Gao G, Fu B (2016) Stemflow of a xerophytic shrub (Salix psammophila) in northern China: implication for beneficial branch architecture to produce stemflow. J Hydrol 539:577–588
Zengeya TA, Wilson JR (2020) The status of biological invasions and their management in South Africa in 2019. South African National Biodiversity Institute, Kirstenbosch and DSI-NRF Centre of Excellence for Invasion Biology, Stellenbosch, p 71. https://doi.org/10.5281/zenodo.3947613
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2024 The Author(s)
About this chapter

Cite this chapter
Geißler, K. et al. (2024). Biodiversity and Ecosystem Functions in Southern African Savanna Rangelands: Threats, Impacts and Solutions. In: von Maltitz, G.P., et al. Sustainability of Southern African Ecosystems under Global Change. Ecological Studies, vol 248. Springer, Cham. https://doi.org/10.1007/978-3-031-10948-5_15
Download citation
DOI: https://doi.org/10.1007/978-3-031-10948-5_15
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-10947-8
Online ISBN: 978-3-031-10948-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)