
Transcriptome Analysis of Embryogenic and Non-Embryogenic Callus of Picea Mongolica
College of Forestry, Inner Mongolia Agricultural University, Huhhot 010019, China
*
Author to whom correspondence should be addressed.
Curr. Issues Mol. Biol. 2023, 45(7), 5232-5247; https://doi.org/10.3390/cimb45070332
Submission received: 29 May 2023
/
Revised: 16 June 2023
/
Accepted: 16 June 2023
/
Published: 21 June 2023
(This article belongs to the Section Molecular Plant Sciences)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Picea mongolica is a rare tree species in China, which is of great significance in combating desertification and improving the harsh ecological environment. Due to the low rate of natural regeneration, high mortality, and susceptibility to pests and cold springs, Picea mongolica has gradually become extinct. At present, somatic embryogenesis (SE) is the most effective method of micro-proliferation in conifers, but the induction rate of embryogenic callus (EC) is low, and EC is difficult to differentiate from non-embryonic callus (NEC). Therefore, the EC and NEC of Picea mongolica were compared from the morphology, histological, physiological, and transcriptional levels, respectively. Morphological observation showed that the EC was white and transparent filamentous, while the NEC was compact and brownish-brown lumpy. Histological analyses showed that the NEC cells were large and loosely arranged; the nuclei attached to the edge of the cells were small; the cytoplasm was low; and the cell gap was large and irregular. In the EC, small cells, closely arranged cells, and a large nucleus and nucleolus were observed. Physiological studies showed significant differences in ROS-scavenging enzymes between the EC and NEC. Transcriptome profiling revealed that 13,267 differentially expressed genes (DEGs) were identified, 3682 were up-regulated, and 9585 were down-regulated. In total, 63 GO terms had significant enrichment, 32 DEGs in plant hormone signal transduction pathway were identified, and 502 different transcription factors (TFs) were characterized into 38 TF families. Meanwhile, we identified significant gene expression trends associated with somatic embryo development in plant hormones (AUX/IAA, YUCCA, LEA, etc.), stress (GST, HSP, GLP, etc.), phenylpropanoid metabolism (4CL, HCT, PAL, etc.), and transcription factors (AP2/ERF, MYB, WOX, etc.). In addition, nine genes were chosen for RT-qPCR, and the results were consistent with RNA-Seq data. This study revealed the changes in morphology, histology, physiology, and gene expression in the differentiation of NEC into EC and laid the foundation for finding the key genes to promote EC formation.
1. Introduction
Picea mongolica W. D. Xu, only distributed in the southeast edge of Otindag Sandy Land in Inner Mongolia, China, is a rare tree species with capabilities of cold tolerance, drought tolerance, and sand-burying tolerance [1]. This wood is mainly used for papermaking, furniture, and musical instrument making, and turpentine can be extracted from its bark. At the same time, as a natural protective barrier, the P. mongolica forest plays a very important role in promoting local agricultural production and the ecological environment in North China [2]. However, due to the low rate of natural regeneration, high mortality, and susceptibility to pests and cold springs, P. mongolica is gradually becoming extinct. In addition, due to the huge genome and high heterozygosity of P. mongolica, seed breeding will inevitably lead to the separation of excellent traits [3]. Therefore, the establishment of somatic embryo regeneration and breeding systems in P. mongolica can accelerate the cultivation of seedlings and also lay the foundation for the genetic improvement of modern molecular breeding technology in P. mongolica.
Plant somatic embryogenesis (SE) is a process in which a single cell or tissue directly induces a callus in vitro without fertilization and eventually differentiates into a whole plant under suitable conditions [4]. Due to its relative genetic stability, repeatability, and efficiency, SE has become an important biotechnology tool and has been widely used in germplasm storage [5], high-quality seedling production [6], artificial seeds [7], and molecular and cell engineering breeding [8,9]. In an experiment on the SE induction of conifers, Durzan and Chalupa first observed the embryoid structure of Pinus banksiana in an in vitro culture [10]. Subsequently, among Pinaceae plants, including Pinus strobus, loblolly pine, and Pinus pinaster, somatic embryos were successfully induced from immature zygotic embryos [11,12,13]. In particular, Picea trees, including Picea morrisonicola [14], Picea koraiensis [15], Picea likiangensis [16], Picea abies [17], and Picea rubens [18], have reported successful SE regeneration. In our previous study, we established a complete SE regeneration system for P. mongolica [19] but found that the induction cycle of its embryogenic callus (EC) is long and the induction efficiency is low. Therefore, a further understanding of the molecular mechanism and key genes involved in SE is necessary to shorten the period of somatic embryo regeneration.
Previous studies have revealed that the EC induction process is controlled by complex genetic factors, including plant hormone-related genes, stress-related genes, and transcription factors [20]. The essential stages of SE are characterized by the induction of the expression of numerous plant hormone-related genes, including PIN [21], ARF [22], YUCCA [23], and AUX/IAA genes [24]. Germin-like proteins (GLPs) were specifically expressed at the initial stage of somatic embryo induction and differentiation in Arabidopsis [25], wheat [26], and cotton [27], and studies in conifers showed that LmGER1 was specifically expressed at the root cap of young embryos [28]. Heat Shock Protein 90 (Hsp90), which is involved in plant developmental processes [29,30,31], showed low expression throughout the early stages of the embryo but was strongly expressed during embryo maturation in Arabidopsis [32]. Many GSTs were found in barley, cotton, and soybean to participate in their embryonic development processes [33,34,35]. In addition, several transcription factors are critical in regulating SE, such as WUSCHEL (WUS) [36], LEAFY COTYLEDON (LEC) [37], AGAMOUS-Like (AGL15) [38], and BABY BOOM (BBM) [39]. However, the drivers of EC induction in P. mongolica have not yet been reported.
In this study, the morphology, cytology, and physiology of the non-embryonic callus (NEC) and EC of P. mongolica were observed, and the gene expression profiles were analyzed by RNA-seq technology. The major candidate genes were identified and discussed in the NEC vs. the EC. This will enhance our knowledge of the SE of P. mongolica and provide theoretical guidance towards solving the problem of EC induction in SE.
2. Materials and Methods
2.1. Plant Materials
P. mongolica seeds, collected from Keshiketeng Banner, Chifeng City, Inner Mongolia Autonomous Region, China, were stored in a refrigerator at 4 °C.
2.2. Callus Induction and Stress Treatment
The mature seeds were soaked in clean water for 24 h. After surface sterilization with 2% NaClO for 6–8 min and sterile water, the mature embryos isolated from the seeds were cultured in induction medium (Table S1). After 28 days, the brown part of the radicle was excised and cultured again in the proliferation medium and supplemented with 5% polyethylene glycol (PEG) 6000 and 20 mM NaCl induction medium, respectively, and the proliferation rate of the EC was counted. After 25 days, the EC and NEC were separated and cultured in the proliferation medium, respectively (Table S1). The EC that grew well in the proliferation medium were selected and put into the differentiation medium for embryoid induction (Table S1). All the cultures were maintained in the dark at 25 ± 1 °C.
2.3. Histological Observation
After culturing on the proliferation medium for 25 days, fresh EC and NEC were selected and fixed in FAA. After washing with 50% alcohol three times, the calli were stained with hematoxylin-eosin at 37 °C for three days. After dehydration with a graded ethanol series (70–100%) and transparent treatment with a xylene series (50–100%), the calli were then embedded in paraffin and sliced using a slicer, with each slice being 10 μm thick. Finally, the sections were sealed with glue, and the morphological differences were observed under the light microscope.
2.4. Physiological Determinations
To determine the activity of the antioxidant enzymes, the EC and NEC were taken at 25 days (0.1 g per sample, 3 replicates) and immediately stored at −80 °C until they were used for the physiological assays. In particular, the activity of superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), ascorbate peroxidase (APX), and polyphenol oxidase (PPO) was determined according to standard methods (Solarbio, Beijing, China).
2.5. RNA-seq
The total RNA was extracted from the EC and NEC using TRIzol® Reagent (Invitrogen, CA, USA). The RNA quality was determined using 1% agarose gels and a 2100 Bioanalyzer (Agilent, CA, USA). RNA-Seq libraries were constructed at Beijing Novogene Technology Co., Ltd. (Beijing, China) using the NEBNext® UltraTM RNA Library Prep Kit (NEB, MA, USA) according to the manufacturer’s recommendations. Finally, the library was performed on the HiSeq2500 platform for high-throughput sequencing.
2.6. Identification of Differentially Expressed Genes (DEGs) and Enrichment Analysis
Through CASAVA base recognition, the high-throughput sequencer transforms the image data of the sequenced fragment into sequence data (reads). The file is in the format of fastq, which mainly contains the sequence information of the sequenced fragment and its corresponding sequence quality information. The reads with the adapter, the reads with n, and the low-quality reads were removed. HISAT2 v2.0.5 (https://ccb.jhu.edu/software/hisat2/index.shtml, accessed on 28 May 2023) was used to compare the paired terminal clean reads with the Norway spruce genome (PlantGenIE.org: Home, accessed on 8 July 2020) to obtain mapped reads. FPKM was used to express the gene expression, and DESeq2 software was used for differential expression analysis. The multiple comparison detection p-values described by Benjamin and Yekutieli [40] were used to check the error detection rate FDR [41,42,43]. The DEGs with p < 0.01 and |log2FC| ≧ 1.5 were considered to be significant. Goseq R and KOBAS software were used to enrich and analyze the DEGs, such as GO and KEGG. TBtools software [44] was used to draw a heatmap according to the gene expression level.
2.7. Quantitative Real-Time PCR (qRT-PCR)
qRT-PCR was carried out to verify the reliability of the RNA-seq data, and the EF1α from P. mongolica was used as the reference gene. Specific primers for 11 DEGs related to EC development for qRT-PCR are listed in Table S2. The qRT-PCR was run on the iCycler iQ system (Bio-Rad, CA, USA) using SYBR qPCR Master Mix (Vazyme, Nanjing, China) according to the manufacturer’s instructions. Three biological and technical repetitions were used for each sample. The relative expression of the DEGs from the qRT-PCR was calculated with the 2−ΔΔCT method. GraphPad. Prism 9.0 (Graphpad, CA, USA) was used to draw histograms according to the gene expression level.
3. Results
3.1. Morphological and Histological Analysis of EC and NEC
To obtain the EC and NEC of P. mongolica, mature embryos were stripped from the sterilized seeds and placed on the induction medium (Figure 1A–C). After induction for 6 days, the whole embryo begins to expand; the radicle and cotyledon turn brown and green, respectively (Figure 1D). A callus began to appear at the hypocotyl on the 8th day (Figure 1E). With an increase in the induction time, the callus became larger, and until the 28th day, white filamentous calli grew on the edge or surface of the massive calli (Figure 1F). After subculturing, the white, transparent, filamentous callus began to grow rapidly (Figure 1G). Finally, the NEC and EC were separated and inoculated in the proliferation medium to make them proliferate and grow stably (Figure 1H,I). In addition, the differentiation of EC can produce a large number of embryoids (Figure 1J).
Histological analysis showed that the NEC cells were large cells with a loose cell arrangement without rules, a small nucleus attached to the cell edge, and a large gap junction (Figure 2A), whereas the EC cells had a large nucleus with a close cell arrangement (Figure 2B). The inner layer of the cell was mostly ordinary parenchyma cells, and the EC appeared outside the cell mass, which can develop into somatic embryos. In addition, the EC was produced on the surface or edge of the NEC (Figure 2C). Therefore, we speculate that the SE of P. mongolica is of external origin.
3.2. The Identification, GO Classification, and KEGG Enrichment Analysis of DEGs
A total of 30,681 DEGs were expressed in both the EC and NEC, and 7012 and 2495 DEGs were uniquely expressed in the NEC and EC, respectively (Figure S1). Additionally, 3682 genes were up-regulated and 9585 genes were down-regulated in the EC compared with the NEC (fold change > 1.5; p-value < 0.05; Figure S2). GO enrichment analysis showed that 34, 10, and 19 GO terms were significantly enriched in biological processes, cell components, and molecular functional classification. Among them, 3914 DEGs were classified into biological processes, mainly enriched in chemical processes, oxidative stress, and metal ion transport. In total, 994 DEGs were classified into cellular components, mainly in the cell periphery, cell wall, and external encapsulating structure; a total of 5529 DEGs were classified into molecular functions, mainly in transcriptional regulator activity, DNA-binding transcription factor activity, and antioxidant activity (Figure S3).
A total of seven pathways were significantly enriched, including photosynthesis, phenylpropanoid biosynthesis, plant hormone signal transduction, phenylpropanoid biosynthesis, the plant hormone signal transduction pathway, photosynthesis-antenna proteins, plant-pathogen interaction, starch and sucrose metabolism, and glutathione metabolism (p-value < 0.05). Strikingly, 32 DEGs are annotated into the plant hormone signal transduction pathway, which is the most annotated pathway of differential genes (Figure S4), indicating that deciphering the discrepancies between the EC and NEC was critical to understanding what enables the NEC to obtain pluripotency and further differentiate into EC.
3.3. Identification and Expression Pattern Analysis of Important DEGs in the Plant Hormone Pathway
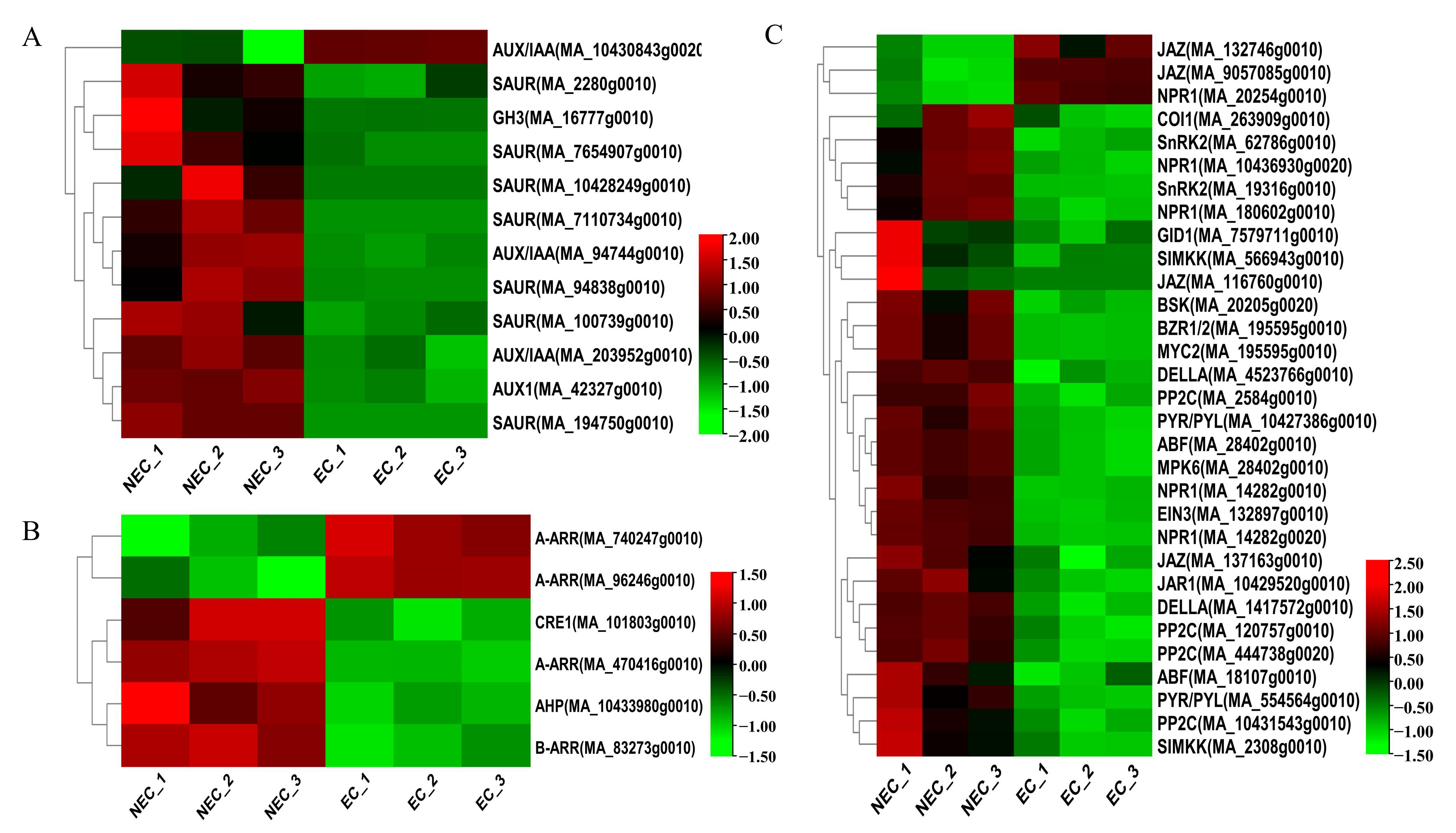
Plant hormone-related genes play a key role in SE [45]. In our study, numerous genes involved in auxin (12 DEGs), cytokinin (6 DEGs), gibberellin (3 DEGs), abscisic acid (10 DEGs), ethylene (4 DEGs), brassinolide (2 DEGs), salicylic acid (5 DEGs), and jasmonic acid (7 DEGs) biosynthesis and signal transduction pathways were differentially expressed in the EC vs. the NEC. For example, the levels of expression of AUX/IAA (MA_10430843g0020) increased from the NEC to EC. Nevertheless, SAUR (MA_100739g0010, MA_194750g0010, MA_7110734g0010, MA_2280g0010, MA_94838g0010, MA_10428249g0010, MA_7654907g0010), AUX1 (MA_42327g0010), and AUX/IAA (MA_203952g0010, MA_94744g0010) were significantly up-regulated (Figure 3A). Cytokinin signal transduction-related DEGs, such as A-ARR (MA_740247g0010, MA_96246g0010), were up-regulated, while CRE1 (MA_101803g0010), AHP (MA_10433980g0010), A-ARR (MA_470416g0010), and B-ARR (MA_83273g0010) were down-regulated (Figure 3B). For jasmonic acid biosynthesis, JAZ (MA_132746g0010, MA_9057085g0010) was up-regulated, while the transcripts, including JAZ (MA_116760g0010, MA_137163g0010), JAR1 (MA_10429520g0010), COI1 (MA_263909g0010), and MYC2 (MA_195595g0010), were down-regulated in the EC vs. NEC. In salicylic acid signal transduction, except for NPR1 (MA_20254g0010), the other four NPR1 genes (MA_10436930g0020, MA_14282g0010, MA_14282g0020, and MA_180602g0010) had low expression in the EC. Gibberellin, abscisic acid, ethylene, and brassinolide signal-related genes decreased from the NEC to EC (Figure 3C). Overall, in the EC vs. the NEC, most of the DEGs had 43 genes that were down-regulated, especially AUX/IAA, SAUR, PP2C, and NPR1, which were 2, 7, 4, and 4, respectively, while 6 genes were up-regulated, of which AUX/IAA, JAZ, NPR1, and A-ARR were 1, 2, 2, and 2, respectively. Therefore, plant hormone-related genes may be the key regulator genes for the EC formation of P. mongolica.
3.4. Identification and Expression Pattern Analysis of Important DEGs in the Phenylpropanoid Metabolism Pathway
Our KEGG enrichment found significant differences in the DEGs between samples in the phenylpropane metabolism pathway (Figure S4). Furthermore, the 8, 1, 1, 5, and 6 DEGs were involved in regulating 4-coumarate CoA ligase (4CL), Shikimate O-hydroxy cinnamoyl transferase (HCT), Scopoletin glucosetransfer (SGT), Cinnamoyl CoA reductase (CCR), and Beta glucose, respectively. However, the 14, 1, 6, 7, and 36 DEGs were down-regulated. Interestingly, the DGEs involved in regulating the levels of phenylalanine amine lyase (PAL), Caffeoyl shikimate esterase (CSE), and Caffeoyl CoA O-methyltransferase (CCoAOMT) were all down-regulated, including 3, 4, and 4, respectively (Figure S5).
3.5. Identification and Expression Profile Analysis of Putative Decisive DEGs Related to Stress Responses
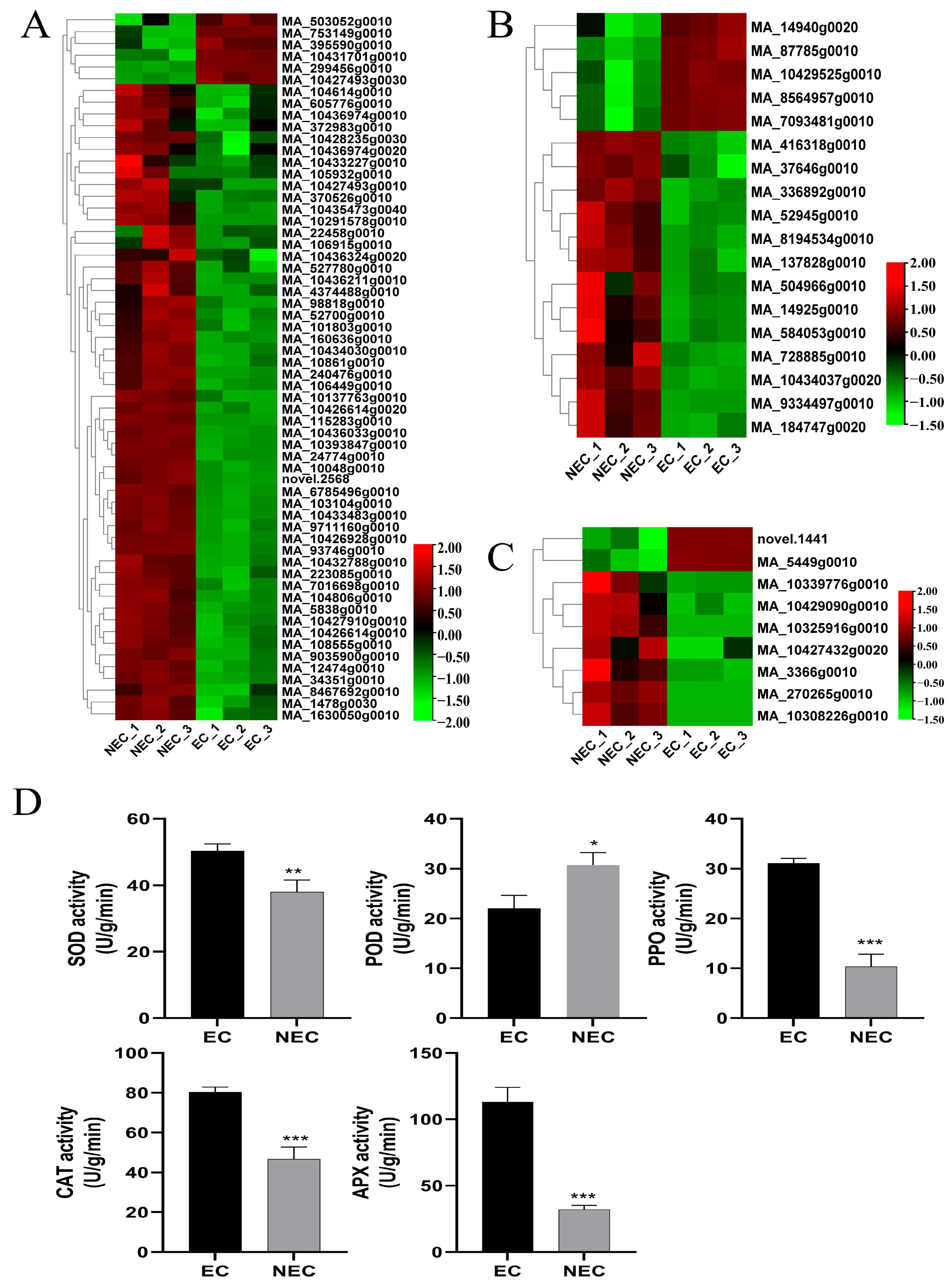
Stress-related genes have been shown to play a vital role during SE [46]. A total of 87 DEGs were identified and analyzed for gene expression, including GST, HSP, and GLP in the EC vs. the NEC (Figure 4A–C). Furthermore, the levels of expression of five GST genes were up-regulated, and MA_7093481g0010 showed the highest up-regulated expression, while 13 GST genes were down-regulated, and MA_10434037g0020 showed the highest down-regulated expression. Moreover, nine GLP genes were identified, of which two were up-regulated and seven were down-regulated. In addition, a large number of HSP genes were identified in the EC vs. the NEC; in total, 5 were up-regulated and 55 were down-regulated. MA_299456g0010 and MA_240476g0010 showed the highest levels of up-regulation and down-regulation, respectively. Meanwhile, the activities of SOD, CAT, PPO, and APX in the EC of P. mongolica were significantly higher than those in the NEC, while the POD activity was higher in the NEC than in the EC (Figure 4D). Among them, the APX enzyme activity of the EC exceeded 100, which was more than three times that of the NEC. These results suggested that the NEC is more susceptible to the physiological factors of browning. Interestingly, we analyzed 100 DEGs enriched in the ROS pathway and found that most of them were related to the antioxidant enzymes POD, SOD, CAT, GPX, and APX (Figure S6). In addition, we conducted drought and salt stress treatments on the callus in Figure 1G and found that appropriate stress can effectively promote EC formation and growth (Figure S7). The above results showed that a large number of stress genes were involved in the expression of EC differentiation in P. mongolica.
3.6. Identification and Expression Profile Analysis of Expression of Decisive Transcription Factors (TFs) Associated with SE
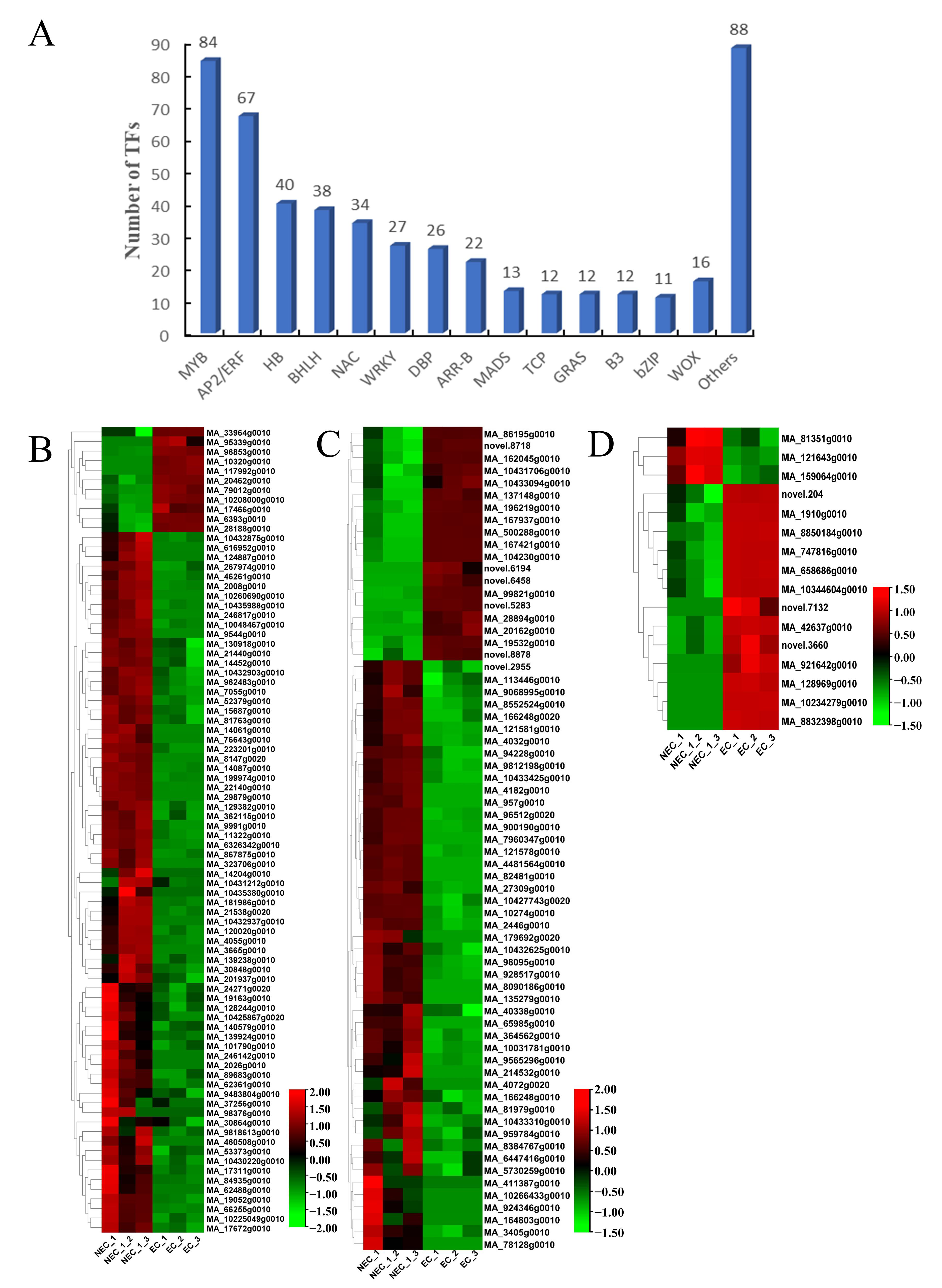
In total, 502 TFs belonging to 38 different TF families were identified in the EC vs. the NEC, mainly in the MYB, AP2/ERF, HB, BHLH, and NAC families (Figure 5A). Furthermore, we analyzed the differential expression levels of the MYB, AP2/ERF, and WOX families in the EC vs. the NEC (Figure 5B–D). The levels of expression of most of the MYB, AP2/ERF, and WOX genes were up-regulated in the EC vs. the NEC. In particular, MYB3 (MA_8147g0020), AP2 (MA_8090186g0010), BBM2 (MA_86195g0010), and WOX4 (MA_121643g0010) had the highest down-regulation expression in the EC vs. the NEC, while MYB305 (MA_10320g0010), ERF071 (MA_196219g0010), and WOX8 (MA_10234279g0010) had the highest up-regulation expression in the EC vs. the NEC. In addition, our results showed 29 down-regulated genes, of which ABI3, ERF, IAA, BRI1, CYP, LEA, YUCCA, ARF, and PIN were 6, 2, 3, 4, 2, 3, 3, 3, and 3, respectively, while 30 genes were up-regulated, of which ABI3, ERF, IAA, BRI1, LEA, ARF, LEC, and PIN were 2, 2, 2, 1, 11, 2, 3, and 7, respectively (Figure S8).
3.7. Validation of the Expression Level of Key Candidate DEGs by qRT-PCR
In order to verify the reliability of the transcriptome data, nine key DEGs involved in SE were analyzed by qRT-PCR (Figure 6). The results showed that all nine DEGs were basically consistent with the transcriptome data, and the correlation between the transcript fold ratios and the qRT-PCR analysis was 0.82, indicating the reliability of the transcriptome data in P. mongolica.
4. Discussion
SE is currently one of the most widely used commercial methods for conifer reproduction, with four stages: EC induction, EC proliferation, SE maturation, and SE germination. EC induction is a critical step for adventitious shoot formation [47]. Many investigations have revealed that certain genes and pathways regulate plant EC induction, with hormone levels being important for EC establishment [20]. In our study, we observed and analyzed the morphological, histological, and physiological differences between the EC and NEC to investigate the mechanisms responsible for the long cycle and low induction efficiency of EC production in P. mongolica. Meanwhile, using RNA-seq, we compared their transcriptional patterns and identified 13,267 DEGs.
To reveal the regulatory mechanism of EC formation, 13,267 DEGs, with 3682 up-regulated and 9585 down-regulated (Figures S1 and S2) genes, were further annotated to plant hormone pathways, particularly the auxin, cytokinin, and ABA pathways, by GO and KEGG analysis (Figures S3 and S4), which is consistent with previous research [48,49], indicating the requirement of plant hormones in SE in P. mongolica. Furthermore, there is no genomic data for P. mongolica, which severely hinders research into its molecular biology. According to our findings, a total of 796.8 M reads were constructed, which will provide basic data support for P. mongolica molecular function research.
Auxin plays a vital role in regulating cell division, differentiation, and the establishment of embryonic polarity [49,50]. YUC1, YUC4, YUC10, and YUC11, which are members of the YUCCAs family that encode for important enzymes in the Trp-dependent auxin production pathway, are expressed in discrete groups of cells during embryogenesis [23].
In addition, many previous studies have shown that GH3 [51], ARF [22], CYP [52], AUX/IAA [24], and SAUR [53] play a key regulatory role during SE. In our study, most of the expression levels of YUCCA, GH3, ARF, IAA, and SAUR, the key genes in Trp synthesis, were drastically down-regulated in the EC vs. the NEC. Only ARF (MA_10432580g0010, MA_2421g0010) and AUX/IAA (MA_10430843g0020, MA_9357208g0010, MA_18664g0010) showed EC specificity with high FPKM (Figure 4A and Figure S4), suggesting that the level of IAA during the NEC was higher than the EC; a low IAA level might promote the NEC to differentiate into the EC. PIN-FORMED1 (PIN1), PIN4, and PIN7 proteins are major factors in the maintenance of auxin gradients in the embryo [54,55]. PIN1 (MA_100472g0010, MA_10430006g0010) and PIN7 (MA_138314g0010) were significantly up-regulated in this study, whereas the seven identified auxin input proteins PIN (MA_57211g0010, novel.7303, novel.477, MA_674082g0010, novel.5169, MA_50550g0010, MA_6089g0010) were significantly down-regulated (Figure S8).
The three cytokinin-encoding receptors CRE1, AHK2, and AHK3 were the first to be discovered in Arabidopsis and are crucial calli-generating factors [56]. On the CRE1 receptor, MA_101803g0010 was identified and was dramatically down-regulated (Figure 3B). The downstream target of AHP is ARR, which may be classified into type A and type B based on the product’s amino acid sequence and expression pattern [57]. By controlling cell cycle-related genes in Arabidopsis, ARR1 can further encourage callus division and growth [58]. In this study, two up-regulated expressions of type A-ARR, one down-regulated expression, and one down-regulated expression of type B-ARR were implicated in the cytokinin signal transduction pathway. The findings above demonstrate that ARR is a critical regulator of P. mongolica’s EC differentiation. Therefore, exogenous cytokinin concentrations can be controlled to hasten EC production during the callus induction of P. mongolica.
Genes involved in the biosynthesis and signal transduction pathways of various hormones, except auxin and cytokinin, as well as gibberellin, ethylene, brassinolide, salicylic acid, jasmonic acid, and abscisic acid were also expressed differently in the NEC and EC (Figure 3C). EC formation and SE germination are impacted by the GA3 content [59]. We discovered that the genes GID1 and DELLA, which are involved in signal transduction and gibberellin production, were up-regulated in the NEC and down-regulated in the EC. SE is promoted by increased ethylene content and the activation of its signal pathway [60]. SIMKK, MPK6, and EIN3, DEGs associated with ethylene signal pathways, were down-regulated in the EC compared to the NEC. The expression of the ethylene-related synthesis genes CTH, cysK, GOT1, EIN2, and EIN3 was, likewise, reduced in the EC in comparison to the NEC in upland cotton [61]. In the EC vs. the NEC, the DEGs of BSK and BZR1/2, which are implicated in BR signal pathways, were down-regulated. The SAR pathway’s main regulatory factor, NPR1 (nonexpressor of pathogenesis-related genes 1), is crucial for plant growth and development [62,63]. Our findings showed that five NPR1 genes were expressed differently in the EC than in the NEC. JAR1, COI1, MYC2, and 2 JAZ were down-regulated in the EC compared to the NEC, but genes linked to JA signal pathways, such as 2 JAZ, were up-regulated. Abscisic acid is associated with SE initiation and expression, and the transition from NEC to EC is followed by a fast increase in endogenous ABA levels [64]. Our results showed that 10 ABA signal pathway-related genes were specifically accumulated in the NEC.
One of the main mechanisms for plant stress adaptation is the phenylpropanoid pathway [65]. This pathway predominates a number of metabolic pathways involved in pathogen defense and abiotic stress, including the formation of flavonoids and lignin [66]. The three phenol propane monomers that make up the majority of lignin are crucial regulators of plant development, cellular mechanical support, and stress response [67]. A crucial enzyme in the production of lignin is phenylalanine amine lyase (PAL), also known as 4-coumarate CoA ligase (4CL) [68]. Additionally, three essential enzymes in the plant shikimate pathway—cinnamoyl CoA reductase (CCR), caffeoyl shikimate ester (CSE), and caffeoyl CoA O-methyltransferase (CCoAOMT)—are involved in the manufacture of significant phenolic acids and are recognized as potent antioxidants [69]. In maize, phenylpropanoid metabolic pathways have several DAPS that are highly abundant in the NEC, according to proteomic analysis of the EC and NEC [70], whereas, in avocado, the phenylpropanoid metabolism pathways accumulate more in the EC than in the NEC [71]. According to our research, the majority of these genes are down-regulated in the EC (Figure S5), suggesting that the NEC needs more lignin.
Stress factors play an important role in somatic embryogenesis. Many studies have proved that stress treatment, including ABA treatment [72], starvation treatment [73], osmotic stress [74], and high-temperature treatment [75], can promote SE and development. In our study, osmotic stress with 20 mM NaCl and 5% PEG6000 boosted the rate of EC proliferation (Figure S7). We measured the enzyme activities of the EC and NEC and discovered that the EC had much greater SOD, PPO, CAT, and APX enzyme activity than the NEC, but the NEC had significantly higher POD enzyme activity than the EC (Figure 4D). Additionally, several SOD, POD, APX, CAT, and GPX genes had distinct levels of expression in the EC compared to the NEC, with the majority of CAT and SOD genes being highly expressed in the EC and the majority of POD genes being highly expressed in the NEC (Figure S6). Previous studies on Gossypium hirsutum L. [76], Agapanthus praecox [49], Araucaria angustifolia [77], Musa spp. [78], and Medicago arborea L. [79] also demonstrated the critical function that antioxidant enzymes play in the development of somatic embryos. The crucial enzyme glutathione-S-transferase (GST), which can also contribute to the metabolism of plant hormones, protects cells by scavenging various stress-induced reactive oxygen species (ROS) [80,81]. The GST gene is essential for giving somatic cells the potential to develop into embryos and can shield plant cells from severe auxin damage [82]. In total, 18 GST genes, of which 5 were up-regulated and 13 were down-regulated, were found to be significantly expressed in the process of cell dedifferentiation by our investigation (Figure 4B). By contributing to the defense of ROS clearance, aiding the somatic embryo in removing too many hazardous substances, boosting cell dedifferentiation, and obtaining embryogenic capacity, GST is triggered by hormone stress. By controlling the cell walls, germination-like protein (GLP) can influence and participate in developing coniferous somatic cells [28]. The expression of the nine GLP genes discovered in this study varied substantially (Figure 4C). Our research, which found 60 heat shock protein genes, is intriguing because it shows that a variety of stressors influence callus development during callus formation and that stress-related genes are essential for callus development (Figure 4A). Three HSP genes were found and cloned by Donget et al. in white spruce somatic embryos, and the transcription of two of these genes was mainly concentrated in the EC. However, by the seedling stage, this gene’s expression was essentially undetectable [83].
Previous research has revealed that transcription factor initiation can govern the transition from NEC to EC or SE induction. LAFL, BBM, AGL15, WIND1, and WOX5 are among them [20]. Our work thoroughly examined the transcription factor data of P. mongolica and discovered 502 differentially expressed TFs, the bulk of which belonged to the MYB and AP2/ERF TF families (Figure 5). MYB, for example, is implicated in numerous plant stress responses and promotes SE [84,85]. Furthermore, the ectopic expression of MYB118 and MYB115 stimulated Arabidopsis root callus formation and increased the expression of the SE maker gene LEC1 [72]. Our findings show that 19, 11, 2, 2, 2, 1, 11, 2, 3, and 7 AP2/ERF, MYB, ABI3, ERF, IAA, BRI1, LEA, ARF, LEC, and PIN genes are significantly up-regulated (Figure 6 and Figure S4). Interestingly, only three of the sixteen WOX genes were discovered to be up-regulated in the EC, whereas the other thirteen genes were strongly expressed, implying that the WOX gene is required for SE in P. mongolica. During somatic embryogenesis, the WUS gene in Arabidopsis promotes the change of auxin-dependent vegetative tissue into embryonic tissue [36,86]. WOX2 is employed as an SE marker gene in pine trees [87]. WOX9 expression is highest during the SE of Picea abies and Medicago truncatula [88,89]. Furthermore, research has shown that ABA induces the expression of the LEA gene and that the exogenous injection of ABA causes a considerable up-regulation of LEA expression [90]. Yang et al. [91,92] discovered multiple LEA genes in carrot and Arabidopsis and discovered that after hormone treatment, the expression of LEA genes in embryogenic tissues increased, which can be employed as molecular indicators of SE. Two up-regulated LEA genes (MA_63890g0010, MA_111913g0010) were found to be strongly expressed in the EC but extremely low in the NEC (Figure S8). Thus, the LEA genes were expressed uniquely in the EC and might be employed as marker genes for P. mongolica EC. Furthermore, research has revealed that the LEC gene can boost the accumulation of stored substances and speed up the transition to embryonic cells and that the LEC1 gene is a crucial regulatory component in embryonic development [93]. Two LEC1 genes were found to be overexpressed in our study. We hypothesize that the LEC1 gene promotes the accumulation of significant amounts of nutrients, such as starch, providing energy for embryonic callus formation and that it may play a positive regulatory role in embryonic callus formation.
In summary, embryogenic ability requires a large number of nutrients to provide energy, and multiple transcription factors have been identified for differential expression in EC formation. These results indicate that the formation of a regulatory network of transcription factors controls the formation of embryogenic calluses in P. mongolica.
5. Conclusions
We used RNA-Seq to perform a comparative transcriptome analysis of the EC and NEC derived from P. mongolica. This study identified some DEGs in the EC vs. the NEC and categorized them according to various biological processes that play a key role in the regulation of SE, such as hormone signaling, stress signaling, phenylpropanoid signaling, and transcription factors (Figure 7). The results of the qRT-PCR showed that the LEC, WOX9, and BBM genes and the GST, GLP, SERF, GH3, AP2, and MYB genes were significantly up-regulated and down-regulated in the EC vs. the NEC, respectively. However, we need to further verify the biological functions and molecular regulatory networks of these key genes through genetic transformation. In summary, our analysis of the differences in the gene profiles between the EC and NEC increased our understanding of the underlying SE in P. mongolica and other coniferous species.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/cimb45070332/s1, Figure S1. Venn diagram of DEGs. Figure S2. Volcano plot of DEGs. Figure S3. GO enrichment analysis of DEGs. Figure S4. Scatter plot of KEGG enrichment analysis. Figure S5. Heatmap of the DEGs related to phenylpropanoid metabolism pathway in EC vs NEC. Figure S6. Heatmap of the DEGs related to the antioxidant enzymes in EC vs NEC. Figure S7. The proliferation rate of EC callus growing in induction medium for 5 to 30 days. Figure S8. Heatmap of the DEGs related to TFs annotated to SE in EC vs NEC. Table S1. Medium composition table. Table S2. qRT-PCR primer information.
Author Contributions
Y.W., W.B. and Y.B. performed the experiment. Y.W., M.S. and H.W. analyzed the data. Y.W. wrote and corrected the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Natural Science Foundation of China (31160166) and the Natural Science Foundation of Inner Mongolia Autonomous Region (No. 2022MS03037).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the sequence data were deposited into the NCBI sequence read archive (SRA) under the BioProject study PRJNA974895 (Samples: SAMN35301251-SAMN35301256).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Xu, W.D.; Li, W.D.; Zheng, Y. A study on taxonomy of P. mongolica in inner Mongolia. Bull. Bot. Res. 1994, 14, 59–69. (In Chinese) [Google Scholar]
- Ha, B.R.; Bai, Y.E.; Hou, W.F.; Qing, S. Research progress and prospect of Picea mongolica of species endemic to sandy land. North. Hortic. 2021, 14, 152–157. (In Chinese) [Google Scholar]
- Nystedt, B.; Street, N.R.; Wetterbom, A.; Zuccolo, A.; Lin, Y.C.; Scofield, D.G.; Vezzi, F.; Delhomme, N.; Giacomello, S.; Alexeyenko, A.; et al. The norway spruce genome sequence and conifer genome evolution. Nature 2013, 497, 579–584. [Google Scholar] [CrossRef] [Green Version]
- Mordhorst, A.P.; Toonen, M.A.J.; Vries, S.C.D.; Meinke, D. Plant embryogenesis. Crit. Rev. Plant Sci. 1997, 16, 535–576. [Google Scholar] [CrossRef]
- Yin, M.H.; Hong, S.R. A simple cryopreservation protocol of Dioscorea bulbifera L. embryogenic calli by encapsulation-vitrifcation. Plant Cell Tissue Organ Cult. 2010, 101, 349–358. [Google Scholar]
- Kessel-Domini, A.; Pérez-Brito, D.; Guzmán-Antonio, A.; Barredo-Pool, F.A.; Mijangos-Cortés, J.O.; Iglesias-Andreu, L.G.; Cortés-Velázquez, A.; Canto-Flick, A.; Avilés-Viñas, S.A.; Rodríguez-Llanes, Y.; et al. Indirect somatic embryogenesis: An efficient and genetically reliable clonal propagation system for Ananas comosus L. Merr. Hybrid “MD2”. Agriculture 2022, 12, 713. [Google Scholar] [CrossRef]
- Manjkhola, S.; Dhar, U.; Joshi, M. Organogenesis, embryogenesis, and synthetic seed production in Arnebia euchroma-a critically endangered medicinal plant of the Himalaya. Cell Dev. Biol. Plant 2005, 41, 244–248. [Google Scholar] [CrossRef]
- Li, B.Q.; Feng, C.H.; Hu, L.Y.; Wang, M.R.; Wang, Q.C. Shoot tip culture and cryopreservation for eradication of Apple stem pitting virus (ASPV) and Apple stem grooving virus (ASGV) from apple rootstocks ‘M9’ and ‘M26’. Ann. Appl. Biol. 2016, 168, 142–150. [Google Scholar] [CrossRef]
- Pathi, K.M.; Tula, S.; Tuteja, N. High frequency regeneration via direct somatic embryogenesis and efcient Agrobacterium-mediated genetic transformation of tobacco. Plant Signal Behav. 2013, 8, e24354. [Google Scholar] [CrossRef] [Green Version]
- Durzan, D.J.; Chalupa, V.; Mia, A.J. Growth and metabolism of cells and tissues of jack pine (Pinus banksiana). 1. The establishment and some characteristics of a proliferated callus from jack pine seedlings. Can J. Bot. 1976, 54, 437–445. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Bernier-Cardou, M.; Cyr, D.R.; Sutton, B.C.S. Influence of gelling agents on culture medium gel strength, water availability, tissue water potential, and maturation response in embryogenic cultures of Pinus strobus L. Vitr. Cell Dev. Biol. Plant 2000, 36, 279–286. [Google Scholar] [CrossRef]
- Pullman, G.S.; Namjoshi, K.; Zhang, Y. Somatic embryogenesis in loblolly pine (Pinus taeda L.): Improving culture initiation rates. Plant Cell Rep. 2003, 22, 85–95. [Google Scholar] [CrossRef]
- Lelu-Walter, M.A.; Bernier-Cardou, M.; Klimaszewska, K. Simplified and improved somatic embryogenesis for clonal propagation of Pinus pinaster (Ait.). Plant Cell Rep. 2006, 25, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.K.; Liao, C.K.; Ho, Y.L. Maturation of somatic embryos in two embryogenic cultures of Picea morrisonicola Hayata as affected by alternation of endogenous IAA content. Plant Cell Tiss. Org. 2008, 93, 257–268. [Google Scholar] [CrossRef]
- Li, C.H.; Liu, B.G.; Kim, T.D.; Moon, H.K.; Choi, Y.E. Somatic embryogenesis and plant regeneration in elite genotypes of Picea koraiensis. Plant Biotechnol. Rep. 2008, 2, 259–265. [Google Scholar] [CrossRef]
- Chen, S.Y.; Chen, S.N.; Fang, C.; Wang, Y.B.; Yi, S.J.; Ning, D.L. Somatic embryogensis in mature zygotic embryos of Picea likiangensis. Biologia 2010, 62, 853–858. [Google Scholar] [CrossRef]
- Hakman, I.; Fowke, L.C.; Von-Arnold, S.; Erikssona, T. The development of somatic embryos in tissue cultures initiated from immature embryos of Picea abies (Norway Spruce). Plants 1985, 38, 53–59. [Google Scholar] [CrossRef]
- Harry, I.S.; Thorpe, T.A. Somatic embryogensis and plant regeneration from mature zygotic embryos of red spruce. Int. J. Plant Sci. 1991, 152, 446–452. [Google Scholar]
- Yan, J.; Peng, P.; Duan, G.Z.; Lin, T.; Bai, Y.E. Multiple analyses of various factors affecting the plantlet regeneration of Picea mongolica (H. Q. Wu) W.D. Xu from somatic embryos. Sci. Rep. 2021, 11, 6694. [Google Scholar] [CrossRef]
- Camille, S.; Loïc, L.; Bertrand, D. Genetic and molecular control of somatic embryogenesis. Plants 2021, 10, 1467. [Google Scholar]
- Robert, H.S.; Grones, P.; Stepanova, A.N.; Robles, L.M.; Lokerse, A.S.; Alonso, J.M.; Weijers, D.; Friml, J. Local auxin sources orient the apical-basal axis in Arabidopsis embryos. Curr. Biol. 2013, 23, 2506–2512. [Google Scholar] [CrossRef] [Green Version]
- Wójcikowska, B.; Gaj, M.D. Expression profifiling of AUXIN RESPONSE FACTOR genes during somatic embryogenesis induction in Arabidopsis. Plant Cell Rep. 2017, 36, 843–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Dai, X.; Zhao, Y. Auxin Synthesized by the YUCCA flavin monooxygenases is essential for embryogenesis and leaf formation in Arabidopsis. Plant Cell 2007, 19, 2430–2439. [Google Scholar] [CrossRef] [Green Version]
- Nic-Can, G.I.; Loyola-Vargas, V.M. The role of the auxins during somatic embryogenesis. In Somatic Embryogenesis: Fundamental Aspects and Applications; Springer: Cham, Switzerland, 2016; pp. 171–182. [Google Scholar]
- Membré, N.; Berna, A.; Neutelings, G.; David, A.; David, H.; Staiger, D.; Sáez-Vásquez, J.; Raynal, M.; Delseny, M.; Bernier, F. cDNA sequence, genomic organization and differential expression of three Arabidopsis genes for germin/oxalate oxidase-like proteins. Plant Mol. Biol. 1997, 35, 459–469. [Google Scholar] [CrossRef]
- Thompson, E.W.; Lane, B.G. Relation of protein synthesis in imbibing wheat embryos to the cell-free translational capacities of bulk mRNA from dry and imbibing embryos. J. Bio. Chem. 1980, 255, 5965–5970. [Google Scholar] [CrossRef]
- Kim, H.J.; Pesacreta, T.C.; Triplett, B.A. Cotton-fiber germin-like protein. II: Immunolocalization, purification, and functional analysis. Planta 2004, 218, 525–535. [Google Scholar] [CrossRef]
- Mathieu, M.; Lelu-Walter, M.A.; Blervacq, A.S.; David, H.; Hawkins, S.; Neutelings, G. Germin-like genes are expressed during somatic embryogenesis and early development of conifers. Plant Mol. Biol. 2006, 61, 615–617. [Google Scholar] [CrossRef] [PubMed]
- Marrs, K.A.; Casey, E.S.; Capitant, S.A.; Bouchard, R.A.; Dietrich, P.S.; Mettler, I.J.; Sinibaldi, R.M. Characterization of two maize hsp90 heat shock protein genes and expression during heat shock, embryogenesis and pollen development. Dev. Genet. 1993, 14, 27–41. [Google Scholar] [CrossRef]
- Reddy, R.; Chaudhary, S.; Patil, P.; Krishna, P. The 90 kDa heat shock protein (hsp90) is expressed throughout Brassica napus seed development and germination. Plants 1998, 131, 131–137. [Google Scholar] [CrossRef]
- Haralampidis, K.; Milioni, D.; Rigas, S.; Hatzopoulos, P. Combinatorial interaction of cis elements specifies the expression of the Arabidopsis AtHsp90-1 gene. Plant Physiol. 2002, 129, 1138–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantinos, P.; Konstantinos, K.; Despina, S.; Polydefkis, H. Tight regulation of expression of two Arabidopsis cytosolic Hsp90 genes during embryo development. J. Exp. Bot. 2005, 56, 633–644. [Google Scholar]
- Vrinten, P.L.; Nakamura, T.; Kasha, K.J. Characterization of cDNAs expressed in the early stages of microspore embryogenesis in barley (Hordeum vulgare) L. Plant Mol. Biol. 1999, 41, 455–463. [Google Scholar] [CrossRef]
- Zeng, F.; Zhang, X.; Zhu, L.; Tu, L.; Guo, X.; Nie, Y. Isolation and characterization of genes associated to cotton somatic embryogenesis by suppression subtractive hybridization and macroarray. Plant Mol. Biol. 2006, 60, 167–183. [Google Scholar] [CrossRef]
- Thibaud-Nissen, F.O.; Shealy, R.T.; Khanna, A.; Vodkin, L.O. Clustering of microarray data reveals transcript patterns associated with somatic embryogenesis in soybean. Plant Physiol. 2003, 32, 118–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, J.; Niu, Q.W.; Frugis, G.; Chua, N.H. The WUSCHEL gene promotes vegetative-to-embryonic transition in Arabidopsis. Plant J. 2002, 30, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Lotan, T.; Ohto, M.A.; Yee, K.M.; West, M.; Harada, J.J. Arabidopsis LEAFY COTYLEDON1 is sufficient to induce embryo development in vegetative cells. Cell 1998, 93, 1195–1205. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Zheng, Y.; Ji, H.; Burnie, W.; Perry, S.E. Gene regulation by the AGL15 transcription factor reveals hormone interactions in somatic embryogenesis. Plant Physiol. 2016, 172, 2374–2387. [Google Scholar] [CrossRef] [Green Version]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.M.; Hattori, J.; Liu, C.M.; Lammeren, A.A.M.; Miki, B.L.A.; et al. Ectopic expression of BABY BOOM triggers aconversion from vegetative to embryonic growth. Plant Cell. 2002, 14, 1737–1749. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR:a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Karami, O.; Aghavaisi, B.; Pour, A.M. Molecular aspects of somatic-to-embryogenic transition in plants. J. Chem. Biol. 2009, 2, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Ruggles, A.; Logan, S.; Mazarakis, A.; Tyson, T.; Bates, M.; Grosse, C.; Reed, D.; Li, Z.G.; Grimwood, J.; et al. Comparative transcriptomics of non-embryogenic and embryogenic callus in semi-recalcitrant and non-recalcitrant upland cotton lines. Plants 2021, 10, 1775. [Google Scholar] [CrossRef]
- Li, W.; Wei, L.; Parris, S.; West, M.; Saski, C.A. Transcriptomic profiles of non-embryogenic and embryogenic callus cells in a highly regenerative upland cotton line (Gossypium hirsutum L.). BMC Dev. Biol. 2020, 20, 25. [Google Scholar]
- Ci, H.T.; Li, C.Y.; Aung, T.t.; Wang, S.L.; Yun, C.; Wang, F.; Ren, X.X.; Zhang, X.X. A comparative transcriptome analysis reveals the molecular mechanisms that underlie somatic embryogenesis in Peaonia ostii ‘Fengdan’. Int. J. Mol. Sci. 2022, 23, 10595. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.H.; Dong, Y.; Du, C.M.; Shi, Y.B.; Teng, Y. Transcriptomic and physiological analyses reveal the acquisition of somatic embryogenesis potential in Agapanthus praecox. Sci. Hortic. 2022, 305, 111362. [Google Scholar] [CrossRef]
- Weijers, D.; Schlereth, A.; Ehrismann, J.S. Auxin triggers transient local signaling for cell specification in Arabidopsis embryogenesis. Dev. Cell. 2006, 10, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Xu, X.; Liu, Z.; Zhang, Z.; XuHan, X.; Lin, Y.; Lai, Z. Global scale transcriptome analysis reveals differentially expressed genes involve in early somatic embryogenesis in Dimocarpus longan Lour. BMC Genom. 2020, 21, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.Y.; Xu, X.Q.; Xu, X.P.; Li, Y.; Zhao, P.C.; Chen, X.H.; Shen, X.; Zhang, Z.H.; Chen, Y.K.; Liu, S.C.; et al. Genome-wide identification, evolution analysis of cytochrome P450 monooxygenase multigene family and their expression patterns during the early somatic embryogenesis in Dimocarpus longan Lour. Gene 2022, 826, 146453. [Google Scholar] [CrossRef]
- Wang, Y.L.; He, S.J.; Long, Y.; Zhang, X.L.; Zhang, X.X.; Hu, H.M.; Li, Z.L.; Hou, F.X.; Ge, F.; Gao, S.B.; et al. Genetic variations in ZmSAUR15 contribute to the formation of immature embryo-derived embryonic calluses in maize. Plant J. 2022, 109, 980–991. [Google Scholar] [CrossRef]
- Weijers, D.; Sauer, M.; Meurette, O.; Friml, J.R.; Ljung, K.; Sandberg, G.; Hooykaas, P.; Offringa, R. Maintenance of embryonic auxin distribution for apical-basal patterning by PIN-FORMED-dependent auxin transport in Arabidopsis. Plant Cell. 2005, 17, 2517–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, F.; Liu, H.H.; Duan, C.Y.; Zhang, B.K.; Wei, G.; Zhang, Y.; Li, S. Arabidopsis JANUS Regulates Embryonic Pattern Formation through Pol II-Mediated Transcription of WOX2 and PIN7. iScience 2019, 19, 1179–1188. [Google Scholar] [CrossRef]
- Inoue, T.; Higuchi, M.; Hashimoto, Y.; Seki, M.; Kobayashi, M.; Kato, T. Identification of CRE1 as a cytokinin receptor from Arabidopsis. Nature 2001, 409, 1060–1063. [Google Scholar] [CrossRef]
- Schaller, G.E.; Kieber, J.J.; Shiu, S.H. Two-Component Signaling Elements and Histidyl-Aspartyl Phosphorelays. Arab. Book 2008, 6, e0112. [Google Scholar] [CrossRef] [Green Version]
- Sakai, H.; Honma, T.; Aoyama, T.; Sato, S.; Kato, T.; Tabata, S.; Oka, A. ARR1, a transcription factor for genes immediately responsive to cytokinins. Science 2001, 294, 1519–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igielski, R.; Kępczyńska, E. Gene expression and metabolite profiling of gibberellin biosynthesis during induction of somatic embryogenesis in Medicago truncatula Gaertn. PLoS ONE 2017, 12, e0182055. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.H.; Liu, Y.B.; Bai, B. Establishment of embryonic shoot-root axis is involved in auxin and cytokinin response during Arabidopsis somatic embryogenesis. Front. Plant Sci. 2015, 5, 792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.H.; Zhu, H.G.; Tian, W.G.; Zhu, S.H.; Xiong, X.P.; Sun, Y.Q.; Zhu, Q.H.; Sun, J. De novo transcriptome analysis reveals insights into dynamic homeostasis regulation of somatic embryogenesis in upland cotton (G. hirsutum L.). Plant Mol. Biol. 2016, 92, 279–292. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Sun, T.J.; Ao, K.; Peng, Y.J.; Zhang, Y.X.; Li, X.; Zhang, Y.L. Opposite roles of salicylic acid receptors NPR1 and NPR3/NPR4 in transcriptional regulation of plant immunity. Cell 2018, 173, 1454–1467. [Google Scholar] [CrossRef]
- Huang, P.X.; Dong, Z.; Guo, P.R.; Zhang, X.; Qiu, Y.P.; Li, B.S.; Wang, Y.C.; Guo, H.W. Salicylic acid suppresses apical hook formation via NPR1-mediated repression of EIN3 and EIL1 in Arabidopsis. Plant Cell 2020, 32, 612–629. [Google Scholar] [CrossRef]
- Liu, W.; Wang, C.L.; Shen, X.L.; Liang, H.W.; Wang, Y.B.; He, Z.Q.; Zhang, D.C.; Chen, F.J. Comparative transcriptome analysis highlights the hormone effects on somatic embryogenesis in Catalpa bungei. Plant Reprod. 2019, 32, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Brunetti, C.; Ferdinando, M.D.; Ferrini, F.; Pollastri, S.; Tattini, M. Functional roles of flavonoids in photoprotection: New evidence, lessons from the past. Plant Physiol. Biochem. 2013, 72, 35–45. [Google Scholar] [CrossRef]
- Zhang, C.H.; Ma, T.; Luo, W.C.; Xu, J.M.; Liu, J.Q.; Wan, D.S. Identification of 4CL genes in desert poplars and their changes in expression in response to salt stress. Genes 2015, 6, 901–917. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.P.; Matthews, M.L.; Williams, C.M.; Shi, R.; Yang, C.; Tunlaya-Anukit, S.; Chen, H.C.; Li, Q.; Liu, J.; Lin, C.Y. Improving wood properties for wood utilization through multi-omics integration in lignin biosynthesis. Nat. Commun. 2018, 9, 1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepelley, M.; Mahesh, V.; McCarthy, J.; Rigoreau, M.; Crouzillat, D.; Chabrillange, N.; de Kochko, A.; Campa, C. Characterization, high-resolution mapping and differential expression of three homologous PAL genes in Coffea canephora Pierre (Rubiaceae). Planta 2012, 236, 313–326. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.H.; Ho, C.T. Antioxidant activities of caffeic acid and its related hydroxycinnamic acid compounds. J. Agr. Food Chem. 1997, 45, 2374–2378. [Google Scholar] [CrossRef]
- Liu, B.B.; Shan, X.H.; Wu, Y.; Su, S.Z.; Li, S.; Liu, H.K.; Han, J.Y.; Yuan, Y.P. iTRAQ-Based quantitative proteomic analysis of embryogenic and non-embryogenic calli derived from a Maize (Zea mays L.) inbred line Y423. Int. J. Mol. Sci. 2018, 19, 4004. [Google Scholar] [CrossRef] [Green Version]
- Olivares-García, C.A.; Mata-Rosas, M.; Peña-Montes, C.; Quiroz-Figueroa, F.; Segura-Cabrera, A.; Shannon, L.M.; Loyola-Vargas, V.M.; Monribot-Villanueva, J.L.; Elizalde-Contreras, J.M.; Ibarra-Laclette, E.; et al. Phenylpropanoids are connected to cell wall fortifification and stress tolerance in avocado somatic embryogenesis. Int. J. Mol. Sci. 2020, 21, 5679. [Google Scholar] [CrossRef] [PubMed]
- Langhansova, L.; Konradova, H.; Vanek, T. Polyethylene glycol and abscisic acid improve maturation and regeneration of Panax ginseng somatic embryos. Plant Cell Rep. 2004, 22, 725–730. [Google Scholar] [CrossRef]
- Touraev, A.; Indrianto, A.; Wratschko, I.; Vicente, O.; Heberle-Bors, E. Efficient microspore embryogenesis in wheat (Triticum aestivum L.) induced by starvation at high temperature. Sex Plant Reprod. 1996, 9, 209–215. [Google Scholar] [CrossRef]
- Kiyosue, T.; Kamada, H.; Har, H. Induction of Somatic Embryogenesis by Salt Stress in Carrot. Plant Tissue Cult. Lett. 2010, 6, 162–164. [Google Scholar] [CrossRef] [Green Version]
- Ikeda-Iwai, M.; Umehara, M.; Satoh, S.; Kamada, H. Stress-induced somatic embryogenesis in vegetative tissues of Arabidopsis thaliana. Plant J. 2003, 34, 107–114. [Google Scholar] [CrossRef]
- Zhou, T.; Yang, X.; Kai, G.; Deng, J.W.; Xu, J.; Gao, W.H.; Lindsey, K.; Zhang, X.L. ROS Homeostasis regulates somatic embryogenesis via the regulation of auxin signaling in cotton. Mol. Cell. Proteom. 2016, 15, 2108–2124. [Google Scholar] [CrossRef] [Green Version]
- Leonardo, J.; Dos, S.A.L.W.; Bueno, C.A.; Barbosa, H.R.; Floh, E.I.S. Proteomic analysis and polyamines, ethylene and reactive oxygen species levels of Araucaria angustifolia(Brazilian pine) embryogenic cultures with different embryogenic potential. Tree Physiol. 2014, 34, 94–104. [Google Scholar]
- Ma, L.; Xie, L.; Lin, G.; Jiang, S.; Chen, H.; Li, H.; Takáč, T.; Šamaj, J.; Xu, C. Histological changes and differences in activities of some antioxidant enzymes and hydrogen peroxide content during somatic embryogenesis of Musa AAA cv. Yueyoukang 1. Sci. Hortic. 2012, 144, 87–92. [Google Scholar] [CrossRef]
- Gallego, P.; Martin, L.; Blazquez, A.; Guerra, H.; Villalobos, N. Involvement of peroxidase activity in developing somatic embryos of Medicago arborea L. Identification of an isozyme peroxidase as biochemical marker of somatic embryogenesis. J. Plant Physiol. 2014, 171, 78–84. [Google Scholar] [CrossRef]
- Lin, Y.F.; Xiao, Q.; Hao, Q.W.; Qian, Z.J.; Li, X.X.; Li, P.; Li, H.; Lian, C. Genome-wide identification and functional analysis of the glutathione S-transferase (GST) family in Pomacea canaliculata. Int. J. Biol. Macromol. 2021, 193, 2062–2069. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; Cummins, I.; Cole, D.J.; Edwards, R. Glutathione-mediated detoxification systems in plants. Curr. Opin. Plant Biol. 1998, 1, 258–266. [Google Scholar] [CrossRef]
- Ganesan, M.; Jayabalan, N. Evaluation of haemoglobin (erythrogen):for improved somatic embryogenesis and plant regeneration in cotton (Gossypium hirsutum L. cv. SVPR 2). Plant Cell Rep. 2004, 23, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.Z.; Dunstan, D.I. Expression of abundant mRNAs during somatic embryogenesis of white spruce [Picea glauca (Moench) Voss]. Planta 1996, 199, 459–466. [Google Scholar] [CrossRef]
- Pratyusha, D.S.; Sarada, D.V.L. MYB transcription factors-master regulators of phenylpropanoid biosynthesis and diverse developmental and stress responses. Plant Cell Rep. 2022, 41, 2245–2260. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Niu, Q.W.; Teng, C.; Li, C.; Mu, J.Y.; Chua, N.H.; Zuo, J.R. Overexpression of PGA37/MYB118 and MYB115 promotes vegetative-to-embryonic transition in Arabidopsis. Cell Res. 2009, 19, 224–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoof, H.; Lenhard, M.; Haecker, A.; Mayer, K.F.X.; Jurgens, G.; Laux, T. The stem cell population of Arabidopsis shoot meristems is maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 2000, 100, 635–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassani, S.B.; Trontin, J.F.; Raschke, J.; Zoglauer, K.; Rupps, A. Constitutive overexpression of a conifer WOX2 homolog affects somatic embryo development in pinus pinaster and promotes somatic embryogenesis and organogenesis in Arabidopsis seedlings. Front. Plant Sci. 2022, 13, 838421. [Google Scholar] [CrossRef]
- Palovaara, J.; Hallberg, H.; Stasolla, C.; Hakman, I. Comparative expression pattern analysis of WUSCHEL-related homeobox 2(WOX2) and WOX8/9 in developing seeds and somatic embryos of the gymnosperm Picea abies. New. Phytol. 2010, 188, 122–135. [Google Scholar] [CrossRef]
- Kurdyukov, S.; Song, Y.; Sheahan, M.B.; Rose, R.J. Transcriptional regulation of early embryo development in the model legume Medicago truncatula. Plant Cell Rep. 2014, 33, 349–362. [Google Scholar] [CrossRef] [Green Version]
- Hatzopoulos, P.; Fong, F.; Sung, Z.R. Abscisis acid regulation of DC8 a carrot embryonic gene. Plant Physiol. 1990, 94, 690–695. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Saitou, T.; Komeda, Y.; Harada, H.; Kamada, H. Late embryogenesis abundant protein in Arabidopsis thaliana homologous to carrot ECP31. Physiol Plant. 2010, 98, 661–666. [Google Scholar] [CrossRef]
- Yang, H.; Saitou, T.; Komeda, Y.; Harada, H.; Kamada, H. Arabidopsis thaliana ECP 63 encoding a LEA protein is located in chromosome 4. Gene 1997, 184, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Kwong, L.W.; Yee, K.M.; Pelletier, J.; Lepiniec, L.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEAFY COTYLEDON2 encodes a B3 domain transcription factor that induces embryo development. Proc. Natl. Acad. Sci. USA 2001, 98, 11806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Callus formation from mature zygotic embryos of Picea mongolica. (A) Cones. (B) Maturing seeds. (C–F) Mature somatic embryos were inoculated on induction medium for 0, 6, 9, and 28 days. (G) Proliferation of white filiform callus after subculturing. (H,I) NEC and EC grown on 25 days of proliferation medium. (J) The embryo is produced by differentiation of the embryogenic callus.
Figure 1.
Callus formation from mature zygotic embryos of Picea mongolica. (A) Cones. (B) Maturing seeds. (C–F) Mature somatic embryos were inoculated on induction medium for 0, 6, 9, and 28 days. (G) Proliferation of white filiform callus after subculturing. (H,I) NEC and EC grown on 25 days of proliferation medium. (J) The embryo is produced by differentiation of the embryogenic callus.

Figure 2.
Microstructure observation of the callus of Picea mongolica. (A) NEC. (B) EC. (C) EC grows on the surface of NEC.
Figure 2.
Microstructure observation of the callus of Picea mongolica. (A) NEC. (B) EC. (C) EC grows on the surface of NEC.

Figure 3.
Identification and expression profile of the DEGs annotated to the hormone synthesis and signal pathways. (A) Auxin signal transduction. (B) Cytokinin signal transduction. (C) Pathways of other hormones, such as gibberellin, abscisic acid, ethylene, brassinolide, salicylic acid, and jasmonic acid.
Figure 3.
Identification and expression profile of the DEGs annotated to the hormone synthesis and signal pathways. (A) Auxin signal transduction. (B) Cytokinin signal transduction. (C) Pathways of other hormones, such as gibberellin, abscisic acid, ethylene, brassinolide, salicylic acid, and jasmonic acid.

Figure 4.
Differential analyses of the stress response of the EC and NEC of Picea mongolica. (A) The heatmap of DEGs related to HSP. (B) The heatmap of DEGs related to GST. (C) The heatmap of DEGs related to GLP. (D) Antioxidant enzyme activity (*, p < 0.05, **, p < 0.01, ***, p < 0.001).
Figure 4.
Differential analyses of the stress response of the EC and NEC of Picea mongolica. (A) The heatmap of DEGs related to HSP. (B) The heatmap of DEGs related to GST. (C) The heatmap of DEGs related to GLP. (D) Antioxidant enzyme activity (*, p < 0.05, **, p < 0.01, ***, p < 0.001).

Figure 5.
Identification and expression profile of the TFs annotated to SE in EC vs. NEC. (A) Identification and categorization of TF families in EC vs. NEC. (B) The heatmap of DEGs related to MYB. (C) The heatmap of DEGs related to AP2/ERF. (D) The heatmap of DEGs related to WOX.
Figure 5.
Identification and expression profile of the TFs annotated to SE in EC vs. NEC. (A) Identification and categorization of TF families in EC vs. NEC. (B) The heatmap of DEGs related to MYB. (C) The heatmap of DEGs related to AP2/ERF. (D) The heatmap of DEGs related to WOX.

Figure 6.
Validation of the expression of nine important candidate DEGs by qRT-PCR. Red and green symbols indicate up-regulated and down-regulated expression of transcriptome sequencing results, respectively.
Figure 6.
Validation of the expression of nine important candidate DEGs by qRT-PCR. Red and green symbols indicate up-regulated and down-regulated expression of transcriptome sequencing results, respectively.

Figure 7.
A proposed molecular regulation diagram of TFs, hormones, phenylpropanoid, and stresses in Picea mongolica somatic embryogenesis. Red words or lines indicate up-regulated gene expression, and green words indicate down-regulated gene expression.
Figure 7.
A proposed molecular regulation diagram of TFs, hormones, phenylpropanoid, and stresses in Picea mongolica somatic embryogenesis. Red words or lines indicate up-regulated gene expression, and green words indicate down-regulated gene expression.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Y.; Wang, H.; Bao, W.; Sui, M.; Bai, Y. Transcriptome Analysis of Embryogenic and Non-Embryogenic Callus of Picea Mongolica. Curr. Issues Mol. Biol. 2023, 45, 5232-5247. https://doi.org/10.3390/cimb45070332
AMA Style
Wang Y, Wang H, Bao W, Sui M, Bai Y. Transcriptome Analysis of Embryogenic and Non-Embryogenic Callus of Picea Mongolica. Current Issues in Molecular Biology. 2023; 45(7):5232-5247. https://doi.org/10.3390/cimb45070332
Chicago/Turabian StyleWang, Yaping, Hao Wang, Wenquan Bao, Mingming Sui, and Yu´e Bai. 2023. "Transcriptome Analysis of Embryogenic and Non-Embryogenic Callus of Picea Mongolica" Current Issues in Molecular Biology 45, no. 7: 5232-5247. https://doi.org/10.3390/cimb45070332