Seed Rain and Seedling Establishment of Picea glauca and Abies balsamea after Partial Cutting in Plantations and Natural Stands

1
Chaire de Recherche sur la Forêt Habitée, Département de Biologie, Chimie et Géographie, Université du Québec à Rimouski, 300 Allée des Ursulines, Rimouski, QC G5L 3A1, Canada
2
Collectif Régional de Développement du Bas-Saint-Laurent, 186 Rue Lavoie, Rimouski, QC G5L 5Z1, Canada
*
Author to whom correspondence should be addressed.
Forests 2019, 10(3), 221; https://doi.org/10.3390/f10030221
Submission received: 7 February 2019
/
Revised: 16 February 2019
/
Accepted: 25 February 2019
/
Published: 1 March 2019
(This article belongs to the Section Forest Ecophysiology and Biology)
Abstract
:The conditions for natural regeneration of white spruce (Picea glauca) and balsam fir (Abies balsamea) in 12 natural stands and five plantations containing both species were investigated 9 to 30 years after partial cutting. We estimated seed input on the ground, measured light reaching the understory, and recorded the presence and age of seedlings smaller than 150 cm in height on six different substrates: mineral soil, moss, rotten wood, litterfall, herbaceous, and dead wood. Partial cutting generally prompted the establishment and growth of seedlings. The number of fir and spruce seedlings is always greater in natural stands than in plantations, a trend likely associated with the reduced abundance of suitable substrate for establishment in the latter. White spruce is significantly associated to rotten wood while fir settles on all types of substrates that cover at least 10% of the forest floor. There is a strong relationship between light intensity and the median height of spruce seedlings, but this relationship is non-significant for fir. Seedlings of both species can survive at incident light intensities as low as 3%, but an intensity of 15% or more seems to offer the best growth conditions. The results of this study provide guidelines for successful forest regeneration following partial cuts in spruce-fir stands.
Keywords:
balsam fir; white spruce; seedlings; partial cut; plantation; naturals stands; light; seed rain1. Introduction
The establishment of natural regeneration in forests having undergone partial cutting offers several silvicultural, economic [1], and ecological [2] benefits. It sustains within-stand demographic stability [2,3,4], decreases reforestation costs [1,5], and promotes structural complexity in both age and the diameter structure of tree populations. The rate of seedling establishment varies according to seed producer density [6], the quality of substrates with regard to germination and early growth [7,8], light [9,10], and harvesting [10]. Therefore, logging operations in a sexually mature forest stand can have a big impact on the natural regeneration of valued tree species [7]. First, the remaining trees can produce more viable seeds when their crown is allowed to further develop [11,12,13]. In addition, logging operations notably modify, through woody residues (e.g., crowns, branches and stumps) left on the ground [14], the characteristics of the substrates on which seeds fall and the resulting seedlings eventually grow. This decaying woody debris [15] constitutes one of the most suitable substrates for germination for Norway spruce (Picea abies [16]) and white spruce (Picea glauca [17]). Finally, partial cutting increases the incident light reaching the forest floor, promoting seedling survival and growth [1,11]. Too much shade in the understory causes increased seedling mortality [9], and incident radiation of less than 10% hinders seedling growth for shade-tolerant tree species like balsam fir (Abies balsamea [9,18]) and white spruce (Picea glauca [19]).
Balsam fir and white spruce are two widespread tree species across North America [20]. They co-occur in naturally regenerated stands, and white spruce plantations are frequently colonized by balsam fir, which seeds-in from nearby forests [21]. The natural regeneration of white spruce is much more difficult to achieve than that of balsam fir [7,8,22,23] but the former has a greater value for the forest industry [24]. This is why the regeneration dynamics of both species in plantations and natural stands after partial harvesting need to be further investigated. In this respect, the influence of seed producers, the establishment of substrate, and light conditions on seed germination and seedling development deserve special consideration [25,26]. In addition, these effects have rarely been compared between natural stands and plantations, whose tree species composition includes variable proportions of both balsam fir and white spruce. Therefore, this study aims to investigate, for natural stands and plantations, (i) whether there is a relationship between seed production and seedling density for balsam fir and white spruce, (ii) whether partial cutting prompts the establishment of regeneration, (iii) whether certain substrates favor regeneration more than others, and (iv) whether light has an effect on seedling growth several years after partial cutting.
2. Materials and Methods
2.1. Study Sites
This study was conducted at the interface of the balsam fir-yellow birch and the balsam fir-white birch bioclimatic domains in Bas-Saint-Laurent, Quebec [27]. A total of 17 out of 24 available sites were randomly selected, including 12 natural stands and 5 plantations. Stand age ranged from 37 to 104 years old. Partial cuttings were done between 8 and 23 years ago for the natural stands and 8 to 30 years ago for the plantations (Table 1). The partial cuts varied in intensity, ranging from 14% to 74% of the total number of trees felled. Natural stands were mostly composed of balsam fir (hereafter referred to as fir) and could be intermixed with white spruce (hereafter referred to as spruce) contributing up to 50% of total basal area. Four spruce plantations were monospecific and 1 had fir contributing to 10% of its total basal area; fir seedlings were present in every spruce plantation. All study sites had a site index [28] greater than 9 m at 25 years for spruce [29] and fir [30]. All plantations originated from abandoned farmlands.
2.2. Measurements and Estimations
For each site, we randomly established 2 sampling units (SUs) spaced 30 to 150 m apart, each measuring 200 m2 (20 m × 10 m); each SU was divided into 50 subunits of 4 m2 (2 m × 2 m). All living trees with a diameter at breast height (DBH) > 5.1 cm were identified at the species level, measured (DBH) and mapped using azimuth and distance from a corner of the SU (reference point). In addition, we established a 5-m buffer zone around the perimeter of each SU; in this buffer, we mapped spruce trees and measured their DBH in order to focus on the potential seed trees of this species. The density (n.m−2) of fallen fir and spruce seeds within each 4 m2 subunit was estimated using models developed by Greene [13]. The first model estimates the number of seeds annually produced by each seed tree (Q) as such:
where m is the mass of a spruce (0.002 g) [19] or fir (0.008 g) [20] seed, and B is seed tree’s basal area (m2) at breast height (1.3 m).
Q = 3067m−0.58B0.92
The second model estimates the number of seeds/m2 scattered on the ground as a function of the distance (x) between a given seed tree and the centroid of each subunit:
where Q(x) defines the number of seeds scattered per m2 at a distance x of a seed tree, a is set to 0.25, which corresponds to a forest with medium seed production [13], and f represents the seed’s final velocity: 0.65 m/s for spruce and 0.86 m/s for fir [13]. Using these two models, we estimated for each seed tree the total number of fallen seeds in each 4 m2 subunit. These values were then compared to the number of spruce and fir seedlings counted in each subunit.
Q(x) = aQe−0.15x f 0.82
In all subunits, we estimated the percentage of cover of each of the six seedbed types: mineral soil, moss, litterfall, herbaceous, rotten wood, and deadwood (corresponding to decomposition classes 3 to 5 and 1 to 2, respectively, according to Reference [15]). We then determined the age and the number of spruce and fir seedlings per substrate type and the following height categories: 1–10 cm, 11–20 cm, 21–30 cm, 31–50 cm, 51–100 cm, and 101–150 cm. The average number of seedlings in SU was then transformed in seedling density per m2 and the percentage of SU with presence of seedling is the stocking coefficient [31]. The age of seedlings <50 cm was estimated using terminal bud scars, while annual growth rings were used for seedlings >50 cm. Age allowed us to differentiate seedlings established before versus after a partial cutting.
Photosynthetically active radiation (PAR, µmol photons·m−2·s−1) was measured 60 cm above ground at the center of each of the 1700 subunits using a Beam-Fraction (BF) sensor-3-type device (Sunshine Sensor (John Wood, Peak Design & Development, Winster, Derbyshire, U.K.); Delta-T Devices (Cambridge, England) BF-3 UM-1.0, Data logger; GP1 v2.1) between 09:00 and 17:00 during the months of June, July, and August. These light measurements were associated with that of a reference sensor placed in an open environment located less than 500 m from the sensor located under forest cover in order to calculate percentage of incident light at each measurement location [32].
Finally, natural stand age was assessed by drawing core samples at a height of 30 cm from the ground from three dominant trees in each SU; these trees were those closest to three randomly selected bearings on a compass. Plantation age corresponds to the year of reforestation; years when a partial cut occurred in each stand type were identified using information obtained from the province’s department of forests unpublished database.
3. Statistical Analyses
3.1. Relationship between the Estimated Amount of Dispersed Seeds and the Number of Seedlings
The relationship between the number of fallen seeds and the number of seedlings of each species was determined using simple linear regression for each stand type. Here and in all subsequent analyses where it was required, normality was verified using the Shapiro–Wilk test (p > 0.05) [33] while homoscedasticity and independence of residuals were assessed by comparing the residuals plotted to the predicted values.
3.2. Comparison of Density and Stocking Coefficient of Seedlings
For each stand type (natural or plantation), we tested whether partial cutting had a significant effect on the density of seedlings and the stocking coefficient for each species. Species, period (before vs after cut), and stand type were considered as fixed variables while site was considered as a random variable. We used generalized linear mixed models (GLMM) [34] to compare seedling density and stocking coefficient between species and between stand types. As the raw data did not have a normal distribution even after logarithmic, square root, arcsin or 1/x transformation trials, [35] we tested several model distributions: Poisson, zero inflated Poisson, negative binomial, and zero inflated negative binomial. Using the Akaike information criterion [34], we determined that the negative binomial model was the most parsimonious. When the analysis revealed significant differences between group means, means were compared between periods, between species or between stand types using Tukey’s multiple comparison test.
3.3. Percentage of Seedlings per Substrate Type
For each species in plantations and natural stands, the percentages of total seedlings germinated on each substrate type were established in order to identify the most suitable substrate for seedling establishment using a Pearson chi-square test [36].
3.4. Relationship between Median Seedling Height and Incident Light
The relationships between the median height of fir and spruce seedlings that germinated after the partial cut and the percentage of light transmitted to the center of each of the 1700 subunits was estimated using simple linear regression.
4. Results
4.1. Relationships between the Estimated Number of Dispersed Seeds and the Number of Seedlings
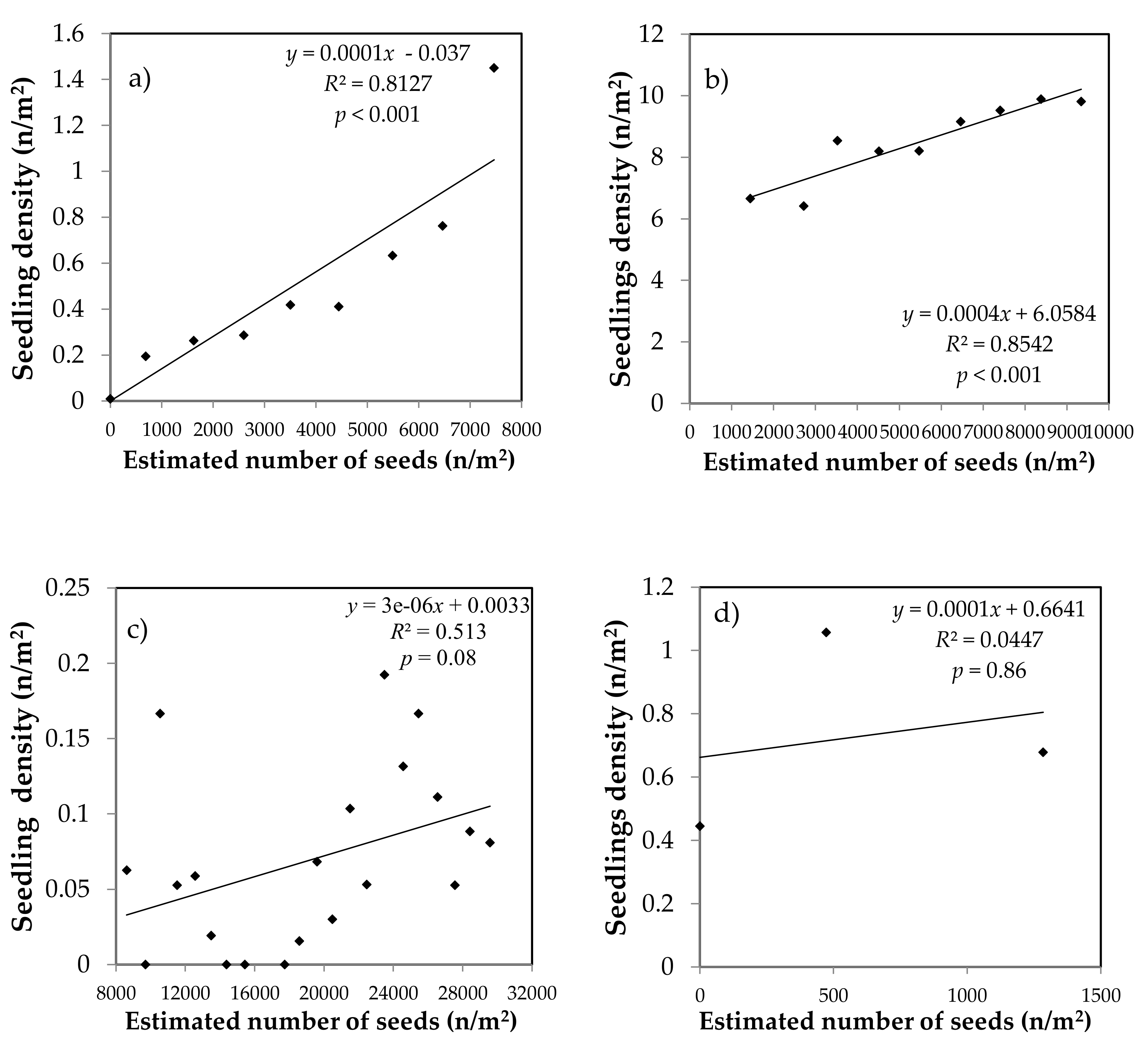
Our results show that there is a close relationship between the estimated number of fallen seeds and the density of fir and spruce seedlings (Figure 1). In the natural stands, the estimated number of dispersed seeds explains 81% and 85% of the variance of fir and spruce seedling density, respectively. For the same amount of seeds, seedling density is 8 to 10 times higher for fir than for spruce. In plantations, the number of fallen seeds explains 51% of the variance of the number of spruce seedlings. According to estimated seed inputs (Figure 1), the establishment of 0.1 spruce seedling m−2 is associated to a seed rain around 30,000 seeds per m2. Although this relationship is not significant for fir, our results suggest that the establishment of a similar density of seedlings would require less than 1000 fir seeds per m2.
4.2. Stocking Coefficient and Density of Seedlings
Partial felling had a significant effect on regeneration in natural stands and plantations. Fir consistently reached a higher stocking coefficient (80%) than spruce (30%) (Figure 2). For fir, there is a significant interaction (p < 0.0001) between treatment and stand type that is associated to an increased stocking coefficient (Figure 2) and density (Figure 3, see Table S1 Supplementary Material) in natural stands and a decrease in plantations. For spruce, partial cuts increase the stocking coefficient and the density of seedlings, especially in natural stands, where both these effects are significant (Figure 2 and Figure 3). Overall, the stocking coefficient and seedlings density of fir and spruce seedlings are significantly higher in natural stands compared to plantations (Figure 2), and fir had a significantly higher seedling density than spruce, both in naturally occurring stands and in plantations (Figure 3).
4.3. Substrate Type Cover and Seedling Establishment in Plantations and Natural Stands
The ground area covered by the various substrate types differs depending on stand origin. The percentages of area covered by litterfall and deadwood are significantly greater (p < 0.05) in plantations than in natural stands, and the opposite is true for moss and rotten wood; herbaceous cover did not differ between natural stands and plantations (Figure 4). In both types of stands, fir and spruce seedlings occurred on all substrate types except deadwood; none were found on mineral soil, as it was absent during the inventory (Figure 4). In natural stands, spruce established more on rotten wood and spruce less on herbaceous substrate than expected by chance (Pearson chi-square test, p < 0.05). Approximately 35% of spruce seedlings were found in the 15% stand area covered by rotten wood and 35% of the seedlings were found in the 40% stand area covered by moss (see Supplementary Materials Figure S1).
4.4. Relationship between the Percentage of Incident Light, Density of Seedlings and Their Height
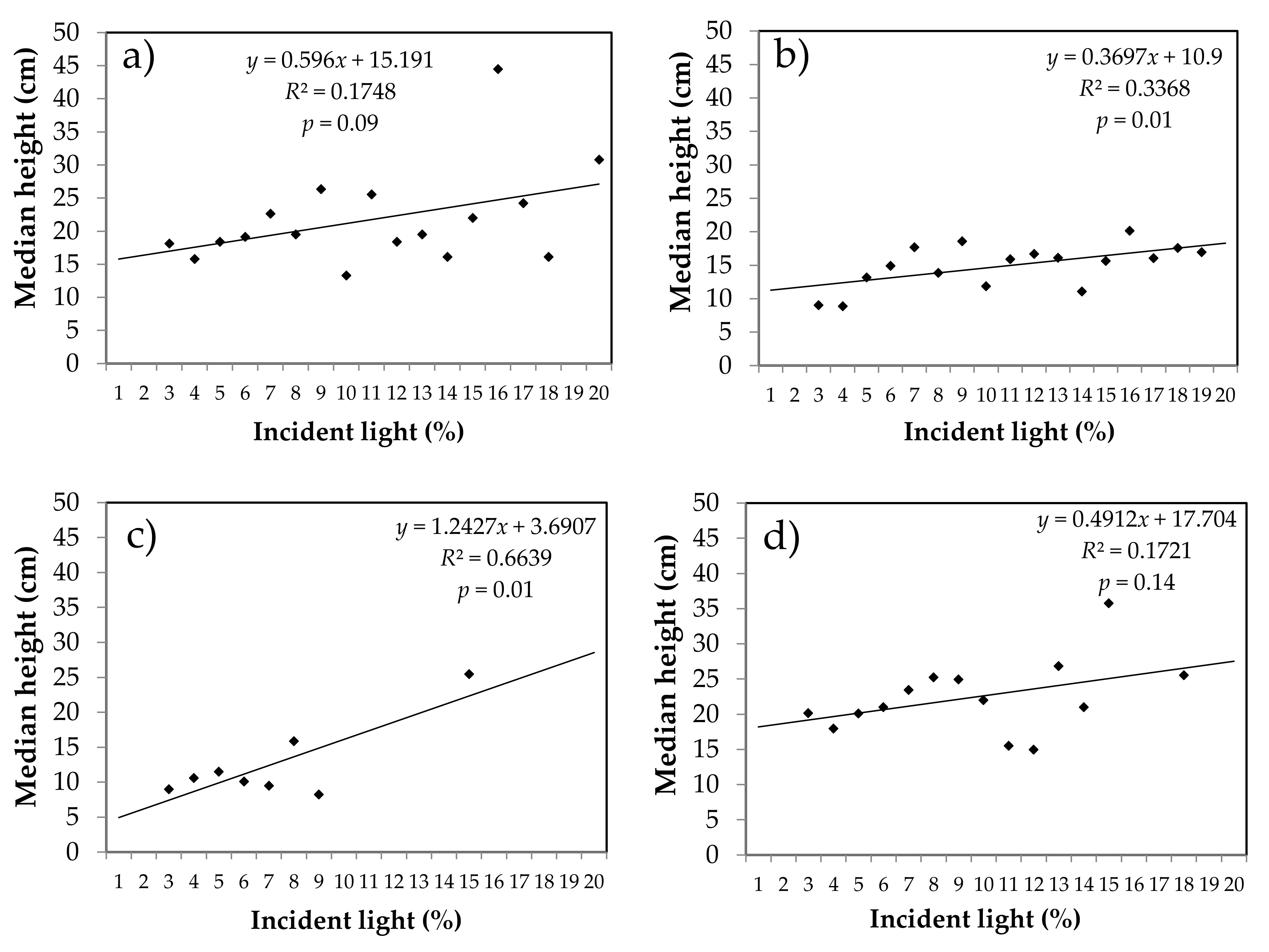
The proportion of incident light measured at 60 cm above ground in partially cut natural stands and plantations reached only 3% to 20% of that measured in the open area. The median height of fir and spruce seedlings in the understory was positively influenced by incident light, which explains 17% of the variance in median height of spruce (p = 0.09) in naturals stands and 66% (p = 0.01) in plantation. For fir, incident light explains 33% of the variance in median height in naturals stands (p = 0.01); in plantations, however, this trend was not significant (p = 0.14) (Figure 5). The relationship between light and the number of seedlings/ha was very weak (R2 = 0.04) but significant (p < 0.05) for fir and spruce in natural stands, while it was not significant (p = 0.62) in plantations.
5. Discussion
This study sheds light on the effects of partial cutting on fir and spruce seedling establishment in natural stands and plantations. Our results suggest that partial cutting promotes seed germination and seedling development. In the northern temperate and boreal forests of Eastern Canada, mainly composed of fir and spruce species, fir seedlings reach densities greater than 35,000 seedlings/ha while spruce only reach 1000 to 3000 seedlings/ha [1,22,39]. In mixed natural stands found elsewhere in the world, the genus Abies is often the one that regenerates most easily when co-occurring with other species [40,41]. Our results indicate that the natural establishment of seedlings is relatively limited in plantations compared to natural stands. This suggests that certain factors favorable to the establishment of regeneration such as substrate type [8,11,23], light [39,42,43], stand structure [44] and seed trees [41] might be more limiting in plantations than in natural stands. The plantations examined in this study were established on abandoned agricultural lands that had been farmed for decades. This presumably modified soil structure by increasing compaction while decreasing aeration and drainage [45,46,47]. In addition, these years of agricultural activity prevented the development of organic soil horizons and reduced the amount of coarse woody debris that littered the ground [48], explaining the reduced rotten wood cover found in our plantations.
The positive influence of rotten wood on natural regeneration from seeds has been reported in several studies [11,23,39]. In this study, rotten wood appears to be mostly associated with the establishment of spruce seedlings, as the latter were found on this substrate in a significantly greater proportion than expected by chance based on substrate relative area (Figure 4 and Figure S1). Our results suggest that a rotten wood cover greater than 15% in natural stands promotes adequate spruce regeneration [49], while it is limited by a low cover (5%) of rotten wood in plantations. A constant supply of woody debris [50] is thus important for the long-term recruitment of rotten wood [17,23,49,51] and the initial survival of spruce seedlings. Fallen rotten wood acts as an ecological filter, catching the smaller spruce seeds and letting many of the larger fir seeds fall on the ground [51]. Rotten wood offers good germination conditions because this substrate has a better water retention capacity than litterfall [7,51,52,53], although several seedlings establish themselves on the latter as it covers a large share of the available area, especially in plantations [54]. There, both species displayed a contrasted ability to survive and grow on this substrate. Field studies suggest that the very short initial roots produced by spruce rarely manage to get through a thick layer of litterfall, as the latter has a coarse porosity and dries up quickly [55,56], inducing a high mortality rate for spruce seedlings [11,57]. Conversely, fir produces longer initial roots [55] able to get through litterfall and reach mineral soil, which makes seedlings less vulnerable to prolonged drought due to its more consistent moisture level [58,59]. This may explains why we can usually find more fir seedlings than spruce seedlings on litterfall, especially in plantations [60]. The herbaceous cover apparently exerts a detrimental effect on spruce establishment, especially in natural stands where this adverse relationship is significant (Figure 4). The slower initial growth of spruce seedlings compared to that of fir [59] would make the former more vulnerable to competitive exclusion by herbs than the latter.
Our results show that the estimated amount of dispersed spruce seeds must be about 45 times greater than that of fir to have an equal number of seedlings of both species in stands where there is a sufficient cover of good settling substrate such as rotten wood. These results are similar to estimations for stands dominated by trembling aspen in which white spruce and balsam fir were the companion species [60]. In plantations where the most suitable substrates for the establishment of spruce only reach 5% cover or less, over 250,000 spruce seeds/m2 would be required to ensure adequate recruitment. Despite the fact that a single white spruce tree can produce up to 250,000 seeds during a mast year [61], such a seed rain is impossible to reach, even in a mature spruce monoculture, where tree density ranges from 1200 to 1500 stems per ha. Thus, rotten wood availability seems to be the limiting factor for spruce regeneration in plantations. Once seeds have germinated, light is important for seedling development [39,43,62,63] and growth [42,64]. According to our results, fir and spruce seedlings can survive for some time at light levels as low as 3% [49,60,64]. Although shade tolerance may decrease as individuals grow from seedling to sapling in some species (e.g. Thuja plicata Donn ex D. Don [65]), but it has been assumed that the minimal incident light requirements for sustained spruce growth is 15% [55]. Fir and spruce seedlings that established themselves after partial cutting do not respond in the same manner to gaps in the canopy [9,66], the growth of fir seedling being less dependent upon variations in percentage of incident light reaching 60 cm above ground compared to spruce. Spruce regeneration may thus be promoted by repeated partial cuts that maintain a minimal light level of 15% in the understory [49].
6. Conclusions
In conclusion, partial cutting promotes seedling establishment in spruce-fir dominated forests, but spruce regeneration remains a challenge, whereas fir readily regenerates in a larger array of conditions [7,22,23]. Following partial cutting in natural stands, a seed input ranging from 7000 to 8000 seeds per m2 of soil surface is required to regenerate one spruce seedling per m2, whereas fir can regenerate ten times more in similar conditions. According to our results, there is a close relationship between seed input and seedling density; in plantations, this relationship lessens for white spruce and is non-significant for fir. The paucity of spruce regeneration despite a high estimated seed input in plantations highlights the limiting role of rotten wood as an establishment substrate for this species. Once seedlings are established, their growth responds positively to incident light, especially for spruce. Therefore, a regime of partial cutting in spruce-fir forests may increase seedling establishment and growth, especially for spruce, provided that cut intensity allows the transmission of 15% incident light to the ground and contributes to maintain a rotten wood ground cover averaging 15%.
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4907/10/3/221/s1. Figure S1: Illustration of the spatial distribution of seed trees, of regeneration and of the rotten wood seedbed substrate in the 200 m2 sampling units, and spatial distribution of white spruce seed trees in the buffer zone of (A) naturals stands and (B) plantations. Table S1: ANOVA table to compare seedling density of each tree type before and after partial cutting and between stand type.
Author Contributions
L.G., L.S. and L.L. conceived and designed the experiment; L.G. analyzed the data, L.S. and L.L. contributed materials and tools, L.G. and L.S. wrote the paper.
Acknowledgments
Funding for this study was provided by the Chaire de recherche sur la forêt habitée of the Université du Québec à Rimouski, the Conférence Régionale des Élus du Bas-Saint-Laurent and the Ministère des Ressources Naturelles et de la Faune du Québec. We would like to thank Nicolas Roy, Philippe Roy for their help with fieldwork and data collection and Kimberly Malcolm for English revision. Special thanks to Alain Caron, Aurélien Nicosia, and Gaetan Daigle for reviewing our statistical analyses.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Olson, M.G.; Meyer, S.R.; Wagner, R.G.; Seymour, R.S. Commercial thinning stimulates natural regeneration in spruce-fir stands. Can. J. For. Res. 2014, 44, 173–181. [Google Scholar] [CrossRef]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for old-growth attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef]
- Lundqvist, L. Stand development in uneven-aged sub-alpine Picea abies stands after partial harvest estimated from repeated surveys. Forestry 2004, 77, 120–129. [Google Scholar] [CrossRef]
- Nilson, K.; Lundqvist, L. Effect of stand structure and density on development of natural regeneration in two Picea abies stands in Sweden. Scand. J. For. Res. 2001, 16, 253–259. [Google Scholar] [CrossRef]
- Lundqvist, L.; Chrimes, D.; Elfving, B.; Mörling, T.; Valinger, E. Stand development after different thinnings in two uneved-aged Picea abies forest in Sweden. For. Ecol. Manag. 2007, 238, 141–146. [Google Scholar] [CrossRef]
- Rive, A.C. Enhancing Natural Regeneration of white Spruce (Picea glauca) via Synchronization of a Mast Year with Site Manipulations in Abitibi, Quebec. Master’s Thesis, Biology Department, Concordia University, Montreal, QC, Canada, 2010. [Google Scholar]
- Calogeropoulos, C.; Greene, D.F.; Messier, C.; Brais, S. The effects of harvest intensity and seedbed type on germination and cumulative survivorship of white spruce and balsam fir in northwestern Quebec. Can. J. For. Res. 2004, 34, 1467–1476. [Google Scholar] [CrossRef]
- Simard, M.-J.; Bergeron, Y.; Sirois, L. Conifer seedling recruitment in a southeastern canadian boreal forest: The importance of substrate. J. Veg. Sci. 1998, 9, 575–582. [Google Scholar] [CrossRef]
- Messier, C.; Doucet, R.; Ruel, J.-C.; Claveau, Y.; Kelly, C.; Lechowicz, M.J. Functional ecology of advance regeneration in relation to light in boreal forests. Can. J. For. Res. 1999, 29, 812–823. [Google Scholar] [CrossRef]
- Blanco, J.A.; Welham, C.; (Hamish) Kimmins, J.P.; Seely, B.; Mailly, D. Guidelines for modeling natural regeneration in boreal forests. For. Chron. 2009, 85, 427–439. [Google Scholar] [CrossRef]
- Gärtner, S.M.; Lieffers, V.J.; Macdonald, S.E. Ecology and management of natural regeneration of white spruce in the boreal forest. Environ. Rev. 2011, 19, 461–478. [Google Scholar] [CrossRef]
- Greene, D.F.; Johnson, E.A. Estimating the mean annual seed production of trees. Ecology 1994, 75, 642647. [Google Scholar] [CrossRef]
- Greene, D.F. Sexual recruitment of trees in strip cuts in eastern Canada. Can. J. For. Res. 2000, 30, 1256–1263. [Google Scholar] [CrossRef]
- Hély, C.; Bergeron, Y.; Flannigan, M.D. Coarse woody debris in the southeastern Canadian boreal forest: Composition and load variations in relation to stand replacement. Can. J. For. Res. 2000, 30, 674–687. [Google Scholar] [CrossRef]
- Christy, E.J.; Mack, R.N. Variation in demography of juvenile Tsuga heterophylla across the substratum mosaic. J. Ecol. 1984, 72, 75–91. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Kalmari, R. Regeneration microsites of Picea abies seedlings in a windthrow area of boreal old-growth forest in southern Finland. Ann. Bot. Fenn. 2003, 40, 401–413. [Google Scholar]
- Simard, M.-J.; Bergeron, Y.; Sirois, L. Substrate and litterfall effects on conifer seedling survivorship in southern boreal stands of Canada. Can. J. For. Res. 2003, 33, 672–681. [Google Scholar] [CrossRef]
- Carswell, F.E.; Doherty, J.E.; Allen, R.B.; Brignall-Theyer, M.E.; Richardson, S.J.; Wiser, S.K. Quantification of the effects of aboveground and belowground competition on growth of seedlings in a conifer– angiosperm forest. For. Ecol. Manag. 2012, 269, 188–196. [Google Scholar] [CrossRef]
- Niensteadt, H.; Zasada, J.C. Picea glauca (Moench) Voss. White spruce. In Silvics of North America; Burns, R.M., Honkola, B.H., Eds.; USDA Forest Service Agriculture Handbook; USDA Forest Service: Washington, DC, USA, 1990; Volume 1, ISBN -13 978-0160271458. [Google Scholar]
- Frank, R.M. Abies balsamea (L.) Mill. Balsam fir. In Silvics of North America; Burns, R.M., Honkola, B.H., Eds.; USDA Forest Service Agriculture Handbook; USDA Forest Service: Washington, DC, USA, 1990; Volume 1, Conifers; pp. 26–35. ISBN -13: 978-0160271458. [Google Scholar]
- Groupe d’experts sur la Sylviculture Intensive de Plantations (GESIP). La Sylviculture Intensive de Plantations dans un Contexte d’aménagement écosystémique; Gouvernement du Québe: Québec City, QC, Canada, 2013; 112p, Rapport du groupe d’experts, sous la direction de M. Barrette et M. Leblanc; Québec; Numéro de publication: DAEF-0354; ISBN 978-2-550-69378-9. [Google Scholar]
- Kneeshaw, D.D.; Bergeron, Y. Ecological factors affecting the abundance of advance regeneration in Quebec’s southwestern boreal forest. Can. J. For. Res. 1996, 26, 888–898. [Google Scholar] [CrossRef]
- Robert, E.; Brais, S.; Harvey, B.D.; Greene, D. Seedling establishment and survival on decaying logs in boreal mixedwood stands following a mast year. Can. J. For. Res. 2012, 42, 1446–1455. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Koubaa, A. Softwoods of Eastern Canada; Special publication 1917-3539; SP-526F FPInnovations Institute: Vancouver, BC, Canada, 2009; ISBN 978-0-86488-538-8. [Google Scholar]
- Klopcic, M.; Bončina, A. Recruitment of tree species in mixed selection and irregular shelterwood forest stands. Ann. For. Sci. 2012, 69, 915–925. [Google Scholar] [CrossRef]
- Schütz, J.P.; Pommerening, A. Can Douglas fir (Pseudotsuga menziesii (Mirb.) Franco.) sustainable grow in complex forest structures? For. Ecol. Manag. 2013, 303, 175–183. [Google Scholar] [CrossRef]
- Saucier, J.-P.; Robitaille, A.; Grondin, P.; Bergeron, J.-F.; Gosselin, J. Les Régions Écologiques du Québec Meridional. (4e version). 2011. Available online: https://mffp.gouv.qc.ca/forets/inventaire/pdf/carte-regions-ecologiques.pdf (accessed on 20 February 2019).
- Alemdag, I.S. National site-index and height growth curves for white spruce growing in natural stands in Canada. Can J. For. Res. 1991, 21, 1466–1474. [Google Scholar] [CrossRef]
- Prégent, G. Tarif de Cubage, Tables de Rendement et Modèles de Croissance pour les Plantations d’épinette Blanche au Québec; Mémoire de recherche forestière No. 160; Ministère des Ressources naturelles et de la Faune, Direction de la recherche forestière, Gouvernement du Québec: Québec City, QC, Canada, 2010; 76p, ISBN 978-2-550-60589-8. [Google Scholar]
- Bédard, S. L’estimation du Potential de Croissance des Stations Forestières: Exemple du Sous-Domaine de la Sapinière à Bouleau Jaune de l’Est du Québec; Mémoire de recherche forestière No. 140; Forêt Québec. Direction de la recherche forestière: Québec City, QC, Canada, 2002; ISBN 2-550-38688-4. [Google Scholar]
- Ministère des Ressources naturelles. Le Guide Sylvicole du Québec—Tome 2—Les concepts et l’application de la Silviculture; Québec, Les Publications du Québec: Québec City, QC, Canada, 2013; 709p.
- Paquette, A.; Bouchard, A.; Cogliastro, A. A less restrictive technique for the estimation of understorey light under variable weather conditions. For. Ecol. Manag. 2007, 242, 800–804. [Google Scholar] [CrossRef]
- Royston, P. Remark AS R94: A remark on Algorithm AS 181: The W test for normality. Appl. Stat. 1995, 44, 547–551. [Google Scholar] [CrossRef]
- Zuur, A.F.; Saveliev, A.A.; Ieno, E.N. Zero Inflated Models and Generalized Linear Mixed Models with R. Newburgh; Highland Statistics Limited: Newburgh, UK, 2012; ISBN 9780957174108. [Google Scholar]
- Chou, Y.M.; Polansky, A.M.M.R.L. Transforming non normal data to normality in statistical process control. J. Qual. Technol. 1998, 30, 2. [Google Scholar] [CrossRef]
- Agresti, A. An Introduction to Categorical Data Analysis, 2nd ed.; John Wiley & Sons: New York, NY, USA, 2007; p. 38. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-97; The R Development Core Team: Vienna, Austria, 2010. [Google Scholar]
- Warnes, G.R.; Bolker, B.; Lumley, T.; Johnson, R.C.; Contributions from Randall C. Johnson Are Copyright. SAIC-Frederick, Inc. Funded by the Intramural Research Program, of the NIH, National Cancer Institute and Center for Cancer Research under NCI Contract NO1-CO-12400. Gmodels: Various R Programming Tools for Model Fitting. 2005 R Package Version 2.15.0. Available online: http://CRAN.Rproject.org/package=gmodels (accessed on 20 February 2019).
- Prévost, M. Effect of cutting intensity on microenvironmental conditions and regeneration dynamics in yellow birch–conifer stands. Can. J. For. Res. 2008, 38, 317–330. [Google Scholar] [CrossRef]
- Cameron, A. Determining the sustainable normal irregular condition: A provisional study on a transformed, irregular mixed species stand in Scotland. Scand. J. For. Res. 2007, 22, 13–21. [Google Scholar] [CrossRef]
- Kuehne, C.; Puettmann, K.J. Natural regeneration in thinned Douglas-stands in western Oregon. J. Sustain. For. 2008, 27, 246–274. [Google Scholar] [CrossRef]
- Carter, G.A.; Smith, W.K. Influence of shoot structure on light interception and photosynthesis in conifers. Plant Physiol. 1985, 79, 1038–1043. [Google Scholar] [CrossRef] [PubMed]
- Lieffers, V.J.; Messier, C.; Stadt, K.J.; Gendron, F.; Comeau, P.G. Predicting and managing light in the understrory of boreal forests. Can. J. For. Res. 1999, 29, 796–811. [Google Scholar] [CrossRef]
- Noguchi, M.; Okuda, S.; Miyamoto, K.; Itou, T.; Inagaki, Y. Composition, size structure and local variation of naturally regenerated broad-leaved tree species in hinoki cypress plantations: A case study in Shikoku, southwestern Japan. Forestry 2011, 84, 493–504. [Google Scholar] [CrossRef]
- Bizzari, L.E.; Collins, C.D.; Brudvig, L.A.; Damschen, E.I. Historical agriculture and contemporary fire frequency alter soil properties in longleaf pine woodlands. For. Ecol. Manag. 2015, 349, 45–54. [Google Scholar] [CrossRef]
- McNabb, D.H.; Startsev, A.D.; Nguyen, H. Soil wetness and traffic level effects on bulk density and airfilled porosity of compacted forest soils. Soil Sci. Soc. Am. J. 2001, 65, 1238–1247. [Google Scholar] [CrossRef]
- Startsev, A.D.; McNabb, D.H. Effects of compaction on aeration and morphology of boreal forest soils in Alberta, Canada. Can. J. Soil Sci. 2009, 89, 45–56. [Google Scholar] [CrossRef]
- McLauchlan, K. The nature and longevity of agricultural impacts on soil carbon and nutrients: A review. Ecosystems 2006, 9, 1364–1382. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Stadt, K.J.; Navratil, S. Age structure and growth of understory white spruce under aspen. Can. J. For. Res. 1996, 26, 1002–1007. [Google Scholar] [CrossRef]
- Leinonen, K.; Leikola, M.; Peltonen, A.; Räsänen, P.K. Kuusen luontainen uudistuminen Pirkka-Hämeen metsälautakunnassa. Summary: Natural regeneration of Norway spruce in Pirkka-Häme Forestry Board District, southern Finland. Acta For. Fenn. 1989, 209, 1–53. [Google Scholar]
- Chećko, E.; Jaroszewicz, B.; Olejniczak, K.; Kwiatkowska-Falińska, A.J. The importance of coarse woody debris for vascular plants in temperate mixed deciduous forests. Can. J. For. Res. 2015, 45, 1154–1163. [Google Scholar] [CrossRef]
- Cornett, M.W.; Reich, P.B.; Puettmann, K.J.; Frelich, L.E. Seedbed and moisture availability determine safe sites for early Thuja occidentalis (Cupressaceae) regeneration. Am. J. Bot. 2000, 87, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Waldron, R.M. Factors Affecting Natural White Spruce Regeneration on Prepared Seedbeds at the Riding Mountain Forest Experimental Area; Government of Canada, Department of Forestry and Rural Development, Forest Research Laboratory: Winnipeg, MB, Canada, 1966. [Google Scholar]
- Novák, J.; Slodičák, M. Structure and accumulation of litterfall under Norway spruce stands in connection with thinnings. J. For. Sci. 2004, 50, 101–108. [Google Scholar] [CrossRef]
- Eis, S. Development of white spruce and alpine fir seedlings on cut-over areas in the central interior of British Columbia. For. Chron. 1965, 41, 419–431. [Google Scholar] [CrossRef]
- Greene, D.F.; Zasada, J.C.; Sirois, L.; Kneeshaw, D.D.; Morin, H.; Charron, I.; Simard, M.-J. A review of the regeneration dynamics of North American boreal forest tree species. Can. J. For. Res. 1999, 29, 824–839. [Google Scholar] [CrossRef]
- Lundqvist, L.; Nilson, K. Regeneration dynamics in an uneven-aged virgin Norway spruce forest in northern Sweden. Scand. J. For. Res. 2007, 22, 304–309. [Google Scholar] [CrossRef]
- Greene, D.F.; Johnson, E.A. Seed mass and early survivorship of tree species in upland clearings and shelterwood. Can. J. For. Res. 1998, 28, 1307–1316. [Google Scholar] [CrossRef]
- Place, I.C.M. The Influence of Seed-Bed Conditions on the Regeneration of Spruce and Balsam Fir; Canadian Department of Northern Affairs and National Resources, Forestry Branch: Gatineau, QC, Canada, 1955. [Google Scholar]
- Wang, G.G.; Kemball, K.J. Balsam fir and white spruce seedling recruitment in response to understory release, seedbed type, and litter exclusion in trembling aspen stands. Can. J. For. Res. 2005, 35, 667–673. [Google Scholar] [CrossRef]
- Hellum, A.K. Grading seed by weight in white spruce. Tree Planters’ Notes 1976, 27, 16–24. [Google Scholar]
- Duchesneau, R.; Morin, H. Early seedling demography in balsam fir seedling banks. Can. J. For. Res. 1999, 29, 1502–1509. [Google Scholar] [CrossRef]
- Dumais, D.; Prévost, M. Management for red spruce conservation in Québec: The importance of some physiological and ecological characteristics—A review. For. Chron. 2007, 83, 378–392. [Google Scholar] [CrossRef]
- Parent, S.; Messier, C. Effets d’un gradient de lumière sur la croissance en hauteur et la morphologie de la cime du sapin baumier régénéré naturellement. Can. J. For. Res. 1995, 25, 878–885. [Google Scholar] [CrossRef]
- Weber, A.; Leckie, S.; Kimmins, J.P.; Gilbert, B.; Blanco, J.A.; Lo, Y.-H. Survival and growth as measures of shade tolerance of planted western redcedar, western hemlock and amabilis fir seedlings in hemlock-fir forests of northern Vancouver Island. For. Ecol. Manag. 2017, 386, 13–21. [Google Scholar] [CrossRef]
- Malcolm, D.C.; Mason, W.L.; Clarke, G.C. The transformation of conifer forests in Britain—Regeneration gap size and silvicultural systems. For. Ecol. Manag. 2001, 151, 7–23. [Google Scholar] [CrossRef]
Figure 1.
Relationship between spruce and fir seed input and seedling density in natural stands (spruce (a) and fir (b)) and plantations (spruce (c) and fir (d)).
Figure 1.
Relationship between spruce and fir seed input and seedling density in natural stands (spruce (a) and fir (b)) and plantations (spruce (c) and fir (d)).

Figure 2.
Stocking coefficient (± standard deviation) of fir (white bar) and spruce (black bar) seedlings before and after partial cut, and total stocking coefficient per species for natural stands (a) and plantations (b) (generalized linear mixed models (glmm)). Lower-case letters indicate a significant difference (p < 0.05) before and after partial cut for a given species in a stand type, while asterisks indicate a significant difference (p < 0.05) between stand types for a given species and period and for total stocking coefficient.
Figure 2.
Stocking coefficient (± standard deviation) of fir (white bar) and spruce (black bar) seedlings before and after partial cut, and total stocking coefficient per species for natural stands (a) and plantations (b) (generalized linear mixed models (glmm)). Lower-case letters indicate a significant difference (p < 0.05) before and after partial cut for a given species in a stand type, while asterisks indicate a significant difference (p < 0.05) between stand types for a given species and period and for total stocking coefficient.

Figure 3.
Seedlings density (± standard deviation (SD)) of fir and spruce before and after partial cut for natural stands (a) and plantations (b) (generalized linear mixed models). Lower-case letters indicate a significant difference (p < 0.05) before vs after partial cut for a given species in each stand type.
Figure 3.
Seedlings density (± standard deviation (SD)) of fir and spruce before and after partial cut for natural stands (a) and plantations (b) (generalized linear mixed models). Lower-case letters indicate a significant difference (p < 0.05) before vs after partial cut for a given species in each stand type.

Figure 4.
Percentage of total seedlings associated with each substrate type and percentage of cover per substrate for natural stands (a) and plantations (b). An asterisk indicates a significant difference (p < 0.05) between a substrate cover and the proportion of seedlings found on this substrate for each species (Pearson chi-square-test). Lower-case letters indicate a significant difference (p < 0.05) in substrate cover between stand types (Pearson chi-square test).
Figure 4.
Percentage of total seedlings associated with each substrate type and percentage of cover per substrate for natural stands (a) and plantations (b). An asterisk indicates a significant difference (p < 0.05) between a substrate cover and the proportion of seedlings found on this substrate for each species (Pearson chi-square-test). Lower-case letters indicate a significant difference (p < 0.05) in substrate cover between stand types (Pearson chi-square test).

Figure 5.
Relationship between median height of spruce and fir seedlings established after the partial cut and the percentage of incident light measured at 60 cm from the ground in natural stands (spruce (a) and fir (b)) and plantations (spruce (c) and fir (d)). n = 858 for fir in naturals stands, n = 243 for fir in plantations, n = 428 for spruce in natural stands, n = 76 for spruce in plantations in a total of 1700 subunits of 4 m2.
Figure 5.
Relationship between median height of spruce and fir seedlings established after the partial cut and the percentage of incident light measured at 60 cm from the ground in natural stands (spruce (a) and fir (b)) and plantations (spruce (c) and fir (d)). n = 858 for fir in naturals stands, n = 243 for fir in plantations, n = 428 for spruce in natural stands, n = 76 for spruce in plantations in a total of 1700 subunits of 4 m2.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Site description in 2009. Standard deviation in parenthesis.
| Origin | Site | Age range of Dominant Tree n = 6 | Years after Partial Cut | Stem Removal | Basal Area (m2/ha) | Mean Diameter at Breast Height (cm) | Stems/ha | Seedlings/ha | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | Spruce | Fir | Spruce | Fir | Spruce | Fir | Spruce | Fir | ||||
| Natural | 1 | 37–40 | 10 | 14.0 | - | 37(0.5) | - | 15(4.1) | − | 1850 | 0 | 4000(4460) |
| 2 | 52–65 | 11 | 67.7 | 3(0) | 26(0.4) | 20 | 16(2.7) | 75(35) | 1325(247) | 875(3833) | 9575(10595) | |
| 3 | 51–71 | 10 | 45.5 | 5(0.5) | 38(0.6) | 20(3.2) | 16(3.4) | 150(141) | 1800(424) | 500(1329) | 4575(6638) | |
| 4 | 46–52 | 9 | 38.2 | 4(0.4) | 27(0.4) | 18(2.6) | 15(3.1) | 125(106) | 1400(247) | 575(1771) | 5300(10470) | |
| 5 | 37–52 | 9 | 36.9 | 16(0.7) | 15(0.7) | 15(4.2) | 17(4.5) | 700(494) | 775(671) | 350(1067) | 6125(6829) | |
| 6 | 48–60 | 9 | 53.0 | 2(0.3) | 25(0.7) | 21(2.3) | 16(4.7) | 75(35) | 1100(212) | 850(3019) | 11,075(16762) | |
| 7 | 55–74 | 10 | 27.9 | 12(1.0) | 26(1.0) | 20(4.7) | 19(5.6) | 300(353) | 875(318) | 2825(5920) | 16,750(16864) | |
| 8 | 62–71 | 9 | 52.1 | 5(1.0) | 36(0.5) | 20(4.7) | 14(4.1) | 150 | 2000 | 550(1161) | 16,850(14575) | |
| 9 | 60–85 | 9 | 11.6 | 7(1.1) | 32(0.8) | 19(6.9) | 16(5.1) | 200(212) | 1350(282) | 550(2089) | 14,425(14190) | |
| 10 | 60–77 | 14 | 73.5 | 4(1.4) | 26(0.7) | 20(6.2) | 19(4.6) | 75(35) | 825(212) | 1625(2667) | 42,050(29602) | |
| 11 | 73–104 | 10 | 74.0 | 8(1.2) | 26(0.7) | 25(4.9) | 18(3.6) | 150(0) | 975(106) | 1225(2809) | 25,950(22551) | |
| 12 | 70–75 | 23 | 60.4 | 7(0.9) | 33(0.7) | 20(5.4) | 18(3.6) | 200(70) | 1225(105) | 200(681) | 3200(8665) | |
| Plantation | 131 | 82 | 30–152 | 42.0 | 29(1.4) | 3(0.3) | 24(5.7) | 10(2.1) | 675(309) | 150(238) | 225(976) | 9350(10000) |
| 14 | 57 | 16–62 | 52.6 | 40(1) | - | 21(5.6) | - | 950(176) | - | 550(1899) | 1650(2664) | |
| 15 | 49 | 8 | 44.3 | 34(0.6) | - | 16(3.3) | - | 1475(247) | - | 0 | 1750(3380) | |
| 16 | 52 | 19−82 | 50.0 | 35(1.4) | - | 22(6.5) | - | 825(196) | - | 0 | 2150(3693) | |
| 17 | 57 | 12 | 65.6 | 29(1.3) | - | 25(6.2) | - | 650(212) | - | 50(351) | 150(596) | |
1 Four sampling units (SUs) were carried out in the same plantation. 2 Two partial cuts in plantation; in these cases, only seedlings established after the last partial cut were considered in the analyses.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gagné, L.; Sirois, L.; Lavoie, L. Seed Rain and Seedling Establishment of Picea glauca and Abies balsamea after Partial Cutting in Plantations and Natural Stands. Forests 2019, 10, 221. https://doi.org/10.3390/f10030221
AMA Style
Gagné L, Sirois L, Lavoie L. Seed Rain and Seedling Establishment of Picea glauca and Abies balsamea after Partial Cutting in Plantations and Natural Stands. Forests. 2019; 10(3):221. https://doi.org/10.3390/f10030221
Chicago/Turabian StyleGagné, Laurent, Luc Sirois, and Luc Lavoie. 2019. "Seed Rain and Seedling Establishment of Picea glauca and Abies balsamea after Partial Cutting in Plantations and Natural Stands" Forests 10, no. 3: 221. https://doi.org/10.3390/f10030221
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.