
Effects of Drought and Flooding on Growth and Physiology of Cinnamomum camphora Seedlings
1
Institute for Forest Resources & Environment of Guizhou, Key Laboratory of Forest Cultivation in Plateau Mountain of Guizhou Province, College of Forestry, Guizhou University, Guiyang 550025, China
2
Jiangxi Provincial Key Laboratory for Restoration of Degraded Ecosystems & Watershed Ecohydrology, Nanchang Institute of Technology, Nanchang 330099, China
3
Research Center of Sichuan Old Revolutionary Areas Development, Sichuan University of Arts and Science, Dazhou 635000, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Forests 2023, 14(7), 1343; https://doi.org/10.3390/f14071343
Submission received: 29 May 2023
/
Revised: 23 June 2023
/
Accepted: 27 June 2023
/
Published: 29 June 2023
(This article belongs to the Special Issue Physiological and Molecular Adaptable Strategies to Abiotic Stress in Forest Trees)
Abstract
:In the context of climate change, future patterns of precipitation are expected to increase the intensity and frequency of drought and flooding stress. This is also likely to result in more frequent flooding–drought alternation events. Drought and flooding stress can have negative effects on tree morphology, physiology, and biochemistry, which can potentially pose long-term threats to tree survival and further disrupt the stability of forest ecosystems. However, it remains unclear how trees would cope with these stressors and their alternations through growth and physiology responses. Potted seedlings of C. camphora were grown under a rainout shelter in the field with four water treatments for 40 days: well-watered (WW), drought stress (DS), flooding stress (FS) and flooding–drought alternation (FDF). Growth, leaf gas exchange, water potential, and biochemical traits were measured. Results show different seedling growth patterns under drought and flooding stress. Drought increased the root-to-shoot ratio (+25%) but flooding favored leaf growth (+33%). Decreases in photosynthesis under drought and flooding were mainly related to stomatal limitations, accompanied by abscisic acid accumulation under drought but not under flooding. Drought reduced water potential, and flooding–drought alternation only decreased water potential in its drought phase. Flooding treatment had no effect on water potential. Drought treatment increased the proline concentrations (+74%) and carbon isotopic composition (+3.7%), but all treatments had no effect on the concentrations of elements in leaves. Drought stress led to carbon depletion in both stem and root, whereas flooding stress primarily induced carbon depletion in the root. Flooding–drought alternation was accompanied by complex physiological responses, including physiological recovery when the stress (flood, drought) was relieved. Our results have shown the different effects of drought and flooding on the growth and physiology of C. camphora seedlings, emphasizing the need to consider specific water stress conditions in future studies and providing a theoretical basis for better management of this tree species in urban areas under variable rainfall patterns.
1. Introduction
The water cycle and atmospheric circulation on Earth may have already been altered by climate change because weather extremes such as severe droughts and floods became more frequent in recent years [1,2]. Drought and flooding stress can have negative effects on tree morphology, physiology, and biochemistry [3,4] and are likely to threaten tree survival in the long run, which may further impact forest carbon-water flux and positive feedback to the atmosphere. Under ongoing climate change, the frequency of drought and flooding is predicted to increase [1]. Particularly, frequent transitions between drought and flooding may do harm to tree species, given they have not evolved a related strategy to acclimate to the rapid changes in water availability. Therefore, an enhanced understanding of how trees cope with drought, flooding, and cycles of these two stressors will provide insights into predicting forest responses to variable rainfall patterns in future climates.
Drought stress has long been observed to affect plant morphology, physiology, and biochemistry. First, drought stress often inhibits growth through the reduction of cell turgor [5]. Under drought stress, plants have the ability to adjust biomass allocation to roots (i.e., increased root-to-shoot ratio), which can explore more resources [6]. Second, drought stress usually reduces leaf water potential, photosynthesis, and stomatal conductance, but can enhance short or long-term leaf-level water-use efficiency [7]. Third, drought stress can have varying effects on biochemistry. For example, drought stress has been found to decrease leaf chlorophyll content but increase proline as an osmolyte for osmotic adjustment and abscisic acid (ABA) for stomatal regulation [8,9]. Moreover, non-structural carbohydrates (NSC) and element stoichiometry can also be changed by drought stress [3].
Previous studies have shown that flooding stress can affect plant morphology, physiology, and biochemistry [10,11,12]. First of all, flooding stress often inhibits plant growth and alters biomass allocation patterns (e.g., reduced root mass and root-to-shoot ratio) [13]. Secondly, flooding stress can reduce leaf photosynthesis, stomatal conductance, and water potential [14,15,16]. Under flooding stress, the decreased hydraulic conductivity and water absorption in the root systems due to root hypoxia further block water transport to leaves. This decline in water transport subsequently reduces leaf water potential and restricts photosynthesis [4,14]. Furthermore, the capacity of roots to absorb nutrients decreases under flooding stress. This can induce nutrient deficiency and decrease chlorophyll content and/or photosynthetic enzymes, which further inhibits photosynthesis [17]. Third, flooding stress also affects proline [18], ABA, and NSC responses [19,20]. For instance, soluble sugars accumulated in leaves under flooding [11,17], functioning as an osmolyte to maintain leaf turgor. In addition, during the post-flood recovery phase, plant physiology may recover from adversity [16]. Trees often undergo complex growth and physiological patterns when subjected to varying water stress. However, few studies have linked the cyclic effects of drought and flooding on tree growth and physiology [21,22], limiting the capacity to predict how trees cope with more variable water availability.
The increased frequency and intensity of drought and flood events have already been a trend in China as well as worldwide [1,23], which may impact tree growth and survival in the future. Alternating cycles of drought and flooding events have increased in China, particularly in the Yangtze River basin and Southwest China. This is primarily due to the influence of East Asian summer monsoon precipitation [24]. Cinnamomum camphora (L.) Presl. is a native tree species in evergreen broadleaved forests in Subtropical China, which is also known as an important species for city landscapes and essential oil production. Consequently, a better understanding of how C. camphora responds to drought, flooding, and their cycles will generate more insights into the capacity of C. camphora to cope with variable rainfall under changing climates, helping with the plantation management of this species. The objective of this study was to address the effects of drought, flooding, and flooding–drought alternation on the growth and physiological responses of C. camphora. We predicted that: (i) drought and flooding stress would decrease total biomass, leaf photosynthesis, stomatal conductance, and water potential, but increase ABA and soluble sugar concentration. Moreover, drought stress would increase the root-to-shoot ratio, but flooding stress would decrease the root-to-shoot ratio; (ii) under flooding–drought alternation conditions, the roots of seedlings would be damaged during periods of flooding, which would lead to a decrease in their resistance to drought. Therefore, it would have a greater negative impact on plant growth and gas exchange compared with drought and flooding alone treatment.
2. Materials and Methods
2.1. Plant Materials and Experimental Design
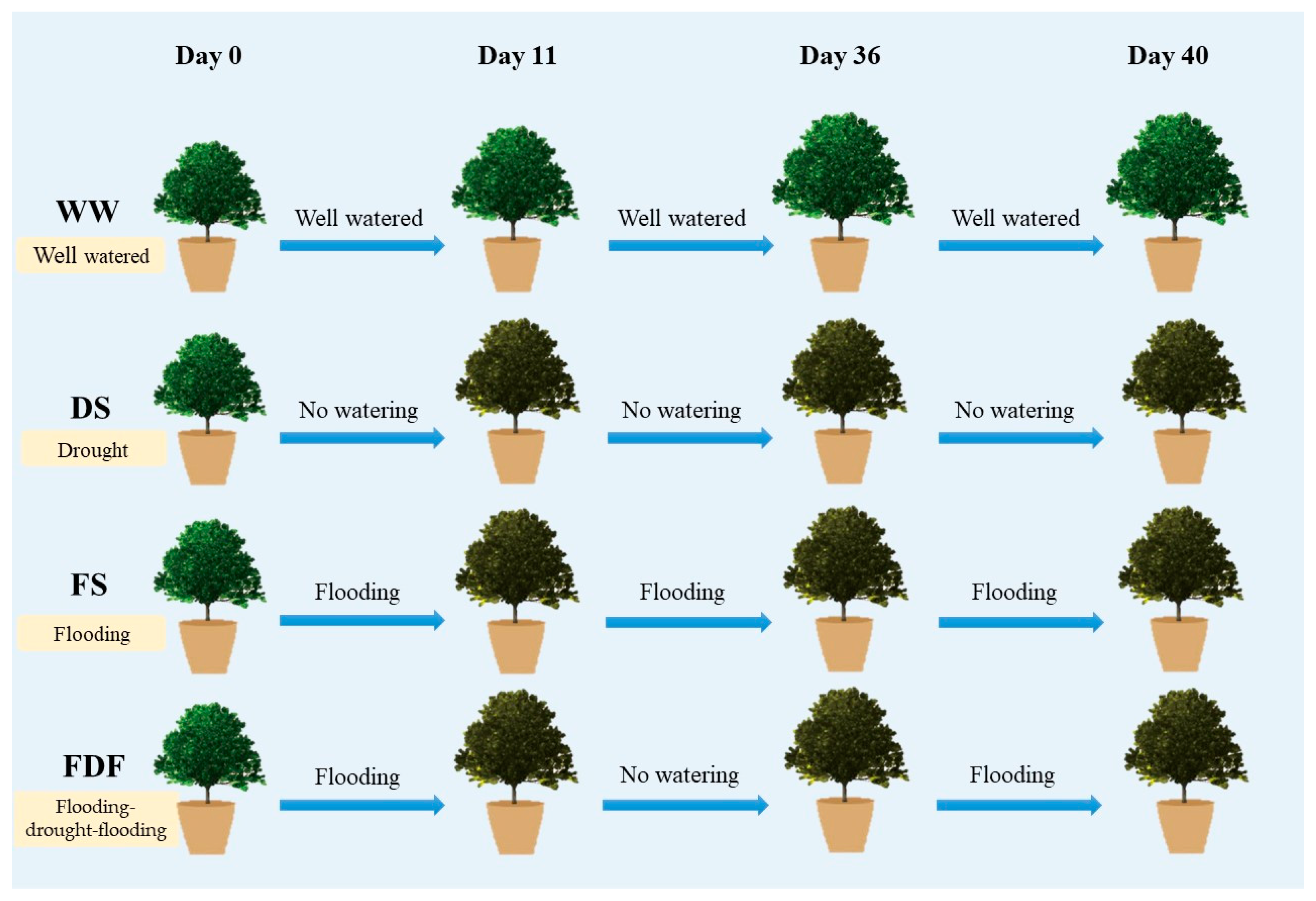
One-year-old C. camphora seedlings were planted into 10 L pots. The soil (red soil; Quaternary Red Earth) used for planting was collected from the top soil layer of the plantation nearby. Pots were randomly placed in a rainout shelter (5 m length × 4 m width × 3 m height) in the field (N 28°41′17.12″, E 116°01′50.16″). Seedlings were well managed with water and weekly application of commercial fertilizer (N ≥ 30 g L−1, P2O5 ≥ 14 g L−1, K2O ≥ 16 g L−1, Fe ≥ 0.14 g L−1 and Mn ≥ 0.06 g L−1) for 4 months prior to treatments. In early September 2017, seedlings with similar heights and diameters were randomly assigned to four water treatments (i.e., no significant differences in height and stem diameter among treatments) (Figure 1): well-watered treatment (WW; n = 10), drought treatment (DS; n = 5), flooding treatment (FS; n = 5) and flooding–drought-flooding treatment (FDF; n = 5). WW seedlings were watered to field capacity throughout the experiment, while DS seedlings were desiccated by withholding water completely for 40 days. Pots of FS seedlings were placed inside 16 L plastic buckets by maintaining the water level 5 cm above the soil surface for 40 days. FDF treatments consisted of three stages: first, seedlings were treated with flooding (same with FS) until stomatal conductance (gs) declined to near 0 mol m−2 s−1 (day 11); seedlings were then moved out from the buckets and left desiccated by withholding water, in order to create the condition for post-flooding recovery (lasting for about 10 days) followed by drought stress; finally, when gs declined to 0 mol m−2 s−1 due to the drought stress (day 36), potted seedlings were treated with another round of flooding towards the end of the experiment (day 40).
2.2. Biomass Measurements
Four seedlings from the WW treatment were destructively harvested as a baseline prior to the experiment, and all the other plants (four seedlings per treatment) were harvested at the end of the experiment. The harvested plants were separated into roots, stems, and leaves, and the roots were thoroughly washed with tap water to remove any soil. Subsequently, all plant organs were immediately oven-dried at 105 °C for 30 min to eliminate any biological activity, followed by further drying at 60 °C for 48 h until a constant weight was achieved. The total biomass (TB) was calculated as the sum of biomass of the root, stem, and leaf. The root-to-shoot ratio (R/S) was determined by dividing root biomass by shoot biomass (leaf and stem biomass). The leaf mass ratio (LMR), stem mass ratio (SMR), and root mass ratio (RMR) were calculated as the ratio of leaf biomass to total biomass, stem biomass to total biomass, and root biomass to total biomass, respectively.
2.3. Gas Exchange and Relative Chlorophyll Content (SPAD) Measurements
During the experiment, we selected mature, fully expanded leaves from 4 seedlings per treatment and conducted leaf gas exchange measurements between 09:00 and 11:00 a.m. Using a portable open path gas exchange system (Licor-6400, Li-Cor, Lincoln, NE, USA) equipped with a red–blue light source (6400-2B). We monitored leaf-saturated light photosynthesis (Asat, µmol m−2 s−1), stomatal conductance (gs, mol m−2 s−1), and transpiration rate (E, mmol m−2 s−1) at light intensity of 1500 µmol m−2 s−1, ambient CO2 of 400 µmol mol−1 and mid-day air temperature (27.9 ± 0.2 °C). The leaf-to-air VPD was 1.57 ± 0.04 kPa. Leaf dark respiration (Rd, µmol m−2 s−1) was measured at night (2 h after sunset) on the same leaf of Asat measurements without light, and at ambient CO2 of 400 µmol mol−1 and air temperature (20 °C). Instantaneous water-use efficiency (WUEi, µmol mol−1) was calculated as Asat/gs.
Leaf SPAD value (relative chlorophyll content, %) was measured on mature, fully expanded leaves from 4 seedlings per treatment using portable chlorophyll system (502, Konica Minolta, Tokyo, Japan) during gas exchange measurements.
2.4. Water Potential Measurements
Pre-dawn leaf water potential (Ψpd, MPa), mid-day leaf water potential (Ψm, MPa), and xylem water potential (Ψx, MPa) were measured from 4 seedlings per treatment using a Scholander-type pressure chamber (PMS 1505D, PMS instruments, Corvalis, OR, USA). In order to ensure the equilibrium of water potential between the soil and plants, seedlings were covered overnight with a plastic film and a foil. Ψpd was determined approximately 1 h before sunrise, while Ψm and Ψx were measured at midday. To determine Ψx, leaves were wrapped with plastic film and foil for at least 1 h before measurements.
2.5. Biochemical Measurements
On day 11, day 36, and day 40, samples were taken from adjacent leaves used for gas exchange measurements from 4 seedlings per treatment (samples were not collected on seedlings of FD and FS treatments on day 36) and frozen in liquid nitrogen prior to biochemical analysis. After extracting samples following Duan et al. [25], abscisic acid (ABA) was measured by Rigol L3000 high-performance liquid chromatography (HPLC) system with a Kromasil C18 reverse phase column (250 mm × 4.6 mm, 5 μm) (Rigol Scientific, Inc., Suzhou, China) [26], while proline was analyzed by the sulfosalicylic acid method [27] and determined colorimetrically at 520 nm. Values of ABA and proline concentration were based on fresh weight.
NSC of oven-dried samples (including starch (ST) and soluble sugars (SS)) was assayed according to the anthrone method [7] at the end of drought and flooding stress. Starch or soluble sugar component ratio (%) was calculated as ST/NSC or SS/NSC.
Total nitrogen (N), total phosphorus (P), and total carbon (C) were assayed with oven-dried samples. Total nitrogen was determined by total organic carbon/total nitrogen analyzer (multi N/C 2100, Jena, Germany) and total phosphorus was determined by molybdenum-antimony resistance colorimetry [28]. Total carbon and leaf carbon isotopic composition (13C, ‰) in oven-dried samples were determined in an element analyzer (NA 2500, Thermo Finnigan, Bremen, Germany) coupled to an isotope ratio mass spectrometer (Delta Plus, Perkin Elmer, Waltham, MA, USA) via a Finnigan ConFloIII interface. All samples for NSC, nutrient, and isotopic composition were collected from harvested seedlings (4 seedlings per treatment) at the end of the experiment.
2.6. Statistical Analyses
Statistical analyses were conducted using SPSS 19.0 (SPSS Inc., Chicago, IL, USA). The time series parameters (i.e., gas exchange, water potential, ABA, proline, SPAD) were analyzed using one-way repeated measures analysis of variance (ANOVA). One-way ANOVA was then assessed to compare means among treatments on each sampling date, followed by Duncan post hoc tests. Similarly, one-way ANOVA was performed on biomass (TB, R/S, RMR, SMR, LMR) and NSC, followed by a Duncan post hoc test for further comparison. The normality and homogeneity were evaluated before all analyses. Significant difference was considered when p < 0.05.
3. Results
3.1. Morphological Responses
At the end of the experiment, compared with the WW treatment, the three water treatments (DS, FS, and FDF) decreased total biomass by 42%, 26%, and 42%, respectively (Table 1), but there was no significant difference among the three treatments. DS treatment significantly increased the root-to-shoot ratio (+25%) and decreased the leaf mass ratio (−30%) relative to WW treatment (p < 0.05), but it did not affect the root mass ratio and stem mass ratio. FS treatment decreased the root-to-shoot ratio (−25%) and root mass ratio (−18%) relative to WW treatment (p < 0.05). Additionally, the leaf mass ratio was higher (+33%). However, FDF treatment only significantly decreased the leaf mass ratio (−41%) compared with WW treatment (p < 0.05). Compared with DS and FDF treatments, the root-to-shoot ratio and root mass ratio of FS treatment were significantly decreased, but the leaf mass ratio was increased. There were no significant differences in root-to-shoot ratio, root mass ratio, and leaf mass ratio between DS and FDF treatments.
3.2. Water Potential
Under DS treatment, leaf water potential was decreased and was lower than WW, FS, and FDF treatments. However, FS treatment did not affect water potential with the exception of Ψx on day 5. FDF treatment significantly decreased water potential compared with WW treatment from day 33 to day 36, with water potential recovered to WW on day 40 (Figure 2).
3.3. Gas Exchange
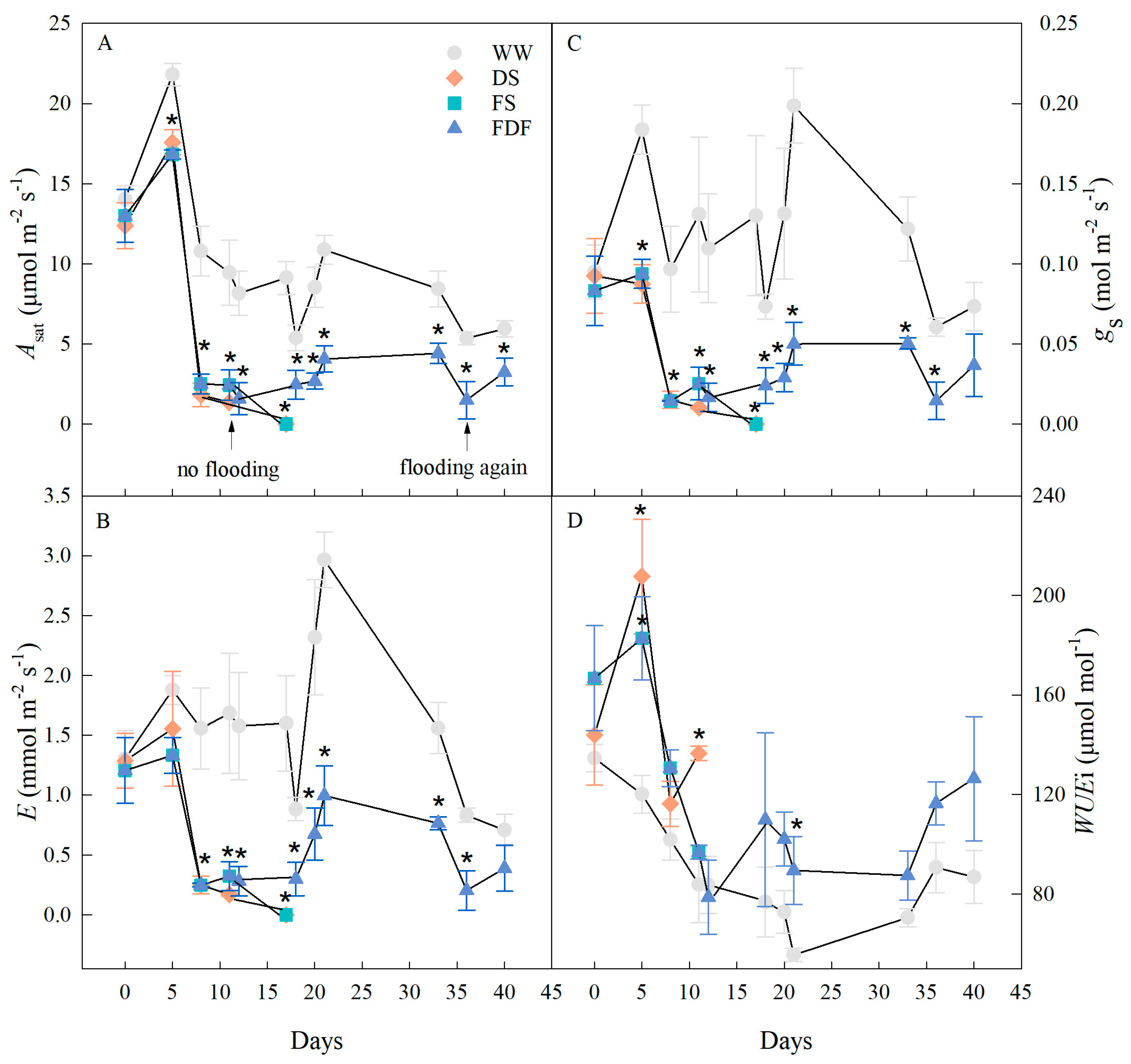
Compared with WW treatment, DS and FS treatments reduced Asat in a similar manner to near zero on day 17. FDF treatment also significantly decreased Asat, while Asat for FDF had a recovery from day 12 to 33 and day 36 to 40 (Figure 3A). In addition, gs and E exhibited similar responses with Asat (Figure 3B,C). We found gs had a negative relationship with ABA irrespective of treatments (Figure 4A). However, Rd did not show a significant difference among treatments (Figure S1), indicating that Rd was less sensitive to water stress than Asat. On day 11, DS, FS, and FDF did not affect SPAD significantly compared with WW treatment, but FDF reduced SPAD on day 36 (−20%) and day 40 (−17%) (Figure S2). Moreover, Asat had a positive relationship with SPAD irrespective of treatments (Figure 4B). Compared with WW treatment, DS treatment enhanced WUEi on days 5 and 11, while FS treatment enhanced WUEi on day 5. FDF treatment increased WUEi only on days 5 and 21 (Figure 3D). Generally, WUEi did not have a relationship with Ψx irrespective of treatments (Figure S3). We also found that Asat, gs, and E had strong correlations with Ψm under DS treatment, but the relationship was not significant under other treatments (WW, FS, and FDF) (Figure 5).
3.4. Non-Structural Carbohydrates
Compared with WW treatment, DS, FS, and FDF treatments did not significantly affect SS and NSC concentration in leaves, but leaf ST concentration was reduced (−51%) in DS treatment and increased (+53%) in FS treatment (Figure 6). In contrast, DS treatment increased the ratio of SS to NSC (+47%) and FS treatment decreased the ratio of SS to NSC (−21%) in leaves (Figure S4). Compared with FS treatment, DS and FDF treatments reduced concentrations of leaf ST (−68%, −42%) and leaf NSC (−35%, −29%), and there was no significant difference between DS and FDF treatments (Figure 6B,C). In addition, the ratio of SS to NSC was ranked as DS > FDF > FS (Figure S4). The effect of DS, FS, and FDF treatments on SS and ST concentration of stem was similar to a leaf. However, FDF treatment increased stem ST concentration (+53%) compared with DS treatment (Figure 6E).
For roots, relative to WW treatment, only DS treatment increased SS concentration (+41%) (Figure 6G), DS and FS decreased ST concentrations of (−68%, −55%) and NSC (−38%, −36%) (Figure 6H,I). Moreover, compared with FDF treatment, DS and FS treatments reduced ST concentration (−73%, −62%). Nevertheless, DS and FS increased the ratio of SS to NSC (+129%, +77%), but FDF treatment did not affect SS, ST, and NSC concentration in the root (Figure 6 and Figure S4).
3.5. ABA and Proline
Compared with the WW treatment, ABA concentration was increased on day 11 in the DS treatment, but it did not change significantly in FS and FDF treatments. Later (days 36 and 40), DS, FS, and FDF treatments did not significantly affect ABA (Figure 7A).
On days 11, 36, and 40, compared with WW, FS, and FDF treatments, DS treatment enhanced proline concentration on day 40, while the proline concentration of FS and FDF treatments did not differ from the WW treatment (Figure 7B).
3.6. Element Stoichiometry and 13C Composition
At the end of the experiment, leaf carbon, nitrogen, and phosphorus stoichiometry (C, N, P, C:N, C:P, and N:P) did not differ significantly among treatments. DS treatment significantly increased 13C compared with WW, FS, and FDF treatments (Table 2), indicating that long-term integrated WUE was enhanced by DS treatment.
4. Discussion
Our study focused on the effects of drought, flooding, and flooding–drought alternation on the growth and physiology of C. camphora seedlings. Consistent with the first hypothesis, our results suggest that drought and flooding stress partially decreased total biomass, photosynthesis, and stomatal conductance. Drought stress increased the root–shoot ratio, but flooding stress decreased the root-to-shoot ratio. Leaf water potential was decreased, and abscisic acid (ABA) was increased only in drought, but soluble sugars in the leaf were not affected by drought and flooding stress. We also found that under flooding–drought alternation conditions, photosynthesis and stomatal conductance showed a tendency of “decrease–rise–decrease–rise”, and gas exchange was decoupled from water potential during the recovery stage. During alternating periods of stress, seedlings had recovery phases, thus there is no evidence to support our second hypothesis that alternating flooding and drought is more detrimental to the growth of C. camphora.
4.1. Plant Biomass and Allocation Responses
In the present study, drought stress had negative impacts on the total biomass of C. camphora seedlings, while it had a positive effect on the root-to-shoot ratio. The result is in line with other studies [6,29], indicating that biomass accumulation and allocation patterns can be altered to acclimate to drought. The increase in root-to-shoot ratio allows plants to absorb more water and nutrients from soil under water-limited conditions [30]. In addition, a reduced leaf mass ratio was found under drought stress. It agrees with findings from previous meta-analysis studies [31], which show that drought can decrease leaf mass ratio to minimize water loss. Plants usually reduce the investment to reproductive parts (i.e., leaf) under drought conditions, in order to explore the water that they need in the soil [32]. In our study, the biomass of C. camphora seedlings was reduced due to flooding stress. This finding is similar to findings in other species, such as Distylium chinense [13] and Magnolia champaca [33], indicating that plant growth is inhibited by flooding stress. Previous studies have shown that flooding reduced the root-to-shoot ratio and root mass ratio [21]. Our study is consistent with the report showing that flooding stress has a greater negative effect on the roots than the stem of Pyrus boissierana [34]. Moreover, in the present study, flooding–drought alternation decreased the total biomass, but it did not affect the root-to-shoot ratio. This is partially consistent with previous research [22] and may be related to the preference for root growth over shoot growth during flooding recovery [35]. Therefore, the increased root-to-shoot ratio during the drought period and its recovery after flooding may offset the decrease in the root-to-shoot ratio during the flooding period.
4.2. Gas Exchange and Water Potential Responses
In this study, leaf and xylem water potentials were decreased under drought stress, which was similar to results from Robinia pseudoacacia seedlings [36]. Moreover, drought stress reduced gs and Asat, which is in line with other results [6,7]. A decrease in stomatal conductance was shown to protect the xylem of leaves and stems from embolism in woody plants [37]. We found that gs had a positive relationship with mid-day leaf water potential, indicating the coupling of stomatal regulation and water status [36]. In addition, it is also found that Asat and gs showed consistent trends, and ABA concentration was increased to regulate stomatal closure under drought stress [38], demonstrating that the reduced Asat was largely due to stomatal limitation. Furthermore, we found a positive correlation between Asat and SPAD in our study, thus the downregulation of Asat was also related to non-stomatal limitation [39], i.e., the degradation of chlorophyll. Previous studies show that drought stress reduced [39], increased [40], or had no effect on chlorophyll content [39]. The variations could be related to the uneven distribution of chlorophyll molecules in leaves, which is affected by the structural organization of the granules in the chloroplast, the number of chloroplasts in the cell, and the number of cells in the tissue [39]. In our study, DS treatment significantly increased the WUEi and δ13C of C.camphor seedlings, indicating that drought stress improved the short- and longer-term water use efficiency of C. camphora seedlings, which is consistent with the results of previous studies [41].
Under flooding stress, leaf water potential can be reduced [14], because water transport could be inhibited by root hypoxia, resulting in a decrease in root hydraulic conductivity and negatively impacting water status in the aboveground part [4]. Some studies have also shown that leaf water potential was increased by flooding [42]. By contrast, in our study, flooding did not significantly affect leaf water potential, indicating that flooding had minimal effects on water transport and water status in the leaves of C. camphora seedlings. In the present study, Asat and gs declined under flooding stress, which agrees with previous reports [15,16]. For example, flooding stress decreased Asat and gs of Populus deltoids [16]. The inhibition of Asat under flooding was also related to stomatal limitation (i.e., reduced gs) and/or non-stomatal limitation (i.e., reduced SPAD) [11,15], and might be due to the accumulation of non-structure carbohydrates in leaves causing feedback inhibition of photosynthesis [15].
Furthermore, Asat and gs have been found to increase during the post-flooding recovery stage [11,16], which is evident under FDF treatment in the present study. Moreover, leaf water potential, Asat, and gs decreased between day 33 to 36 and increased between day 36 to 40, mainly due to drought stress and following recovery, which is consistent with previous reports [7,36], showing that C. camphora has the capacity to recover after drought. We found that SPAD values on days 36 and 40 in the drought stage of FDF treatment were lower than the control, which could partially explain the decline of Asat [39]. Moreover, we found decoupling between leaf gas exchange (i.e., Asat and gs) and water potential under FDF treatment, which is perhaps related to its complex processes (flooding, flooding recovery, and drought recovery). Similarly, some other studies also found that water potential and photosynthesis were decoupled during these processes [17,43].
Collectively, drought, flooding and flooding–drought alternation reduced Asat and gs of C. camphora seedlings, but water potentials changed significantly only during drought stress and flooding–drought alternation (drought stress stage and drought recovery stage). These results indicate that drought stress had a greater influence on the water transport of C. camphora seedlings than flooding stress. Under drought stress, the occurrence of xylem embolism often reduces hydraulic conductivity, leading to lower water transport to leaves and declines in leaf water potential [7]. However, under flooding stress in the present study, water potential was not reduced reflecting that the flooding treatment in this study did not affect water transport.
4.3. NSC, Proline, and Nutrient Responses
The storage of non-structural carbohydrates plays a key role in providing energy reserves for plants, particularly during environmental stress [44]. Our results demonstrate that the decrease in NSC concentration of all organs under drought stress was mainly due to the decrease in starch, implying the imbalance between carbon consumption and carbon availability [45]. Other studies have also shown that under drought stress, the NSC concentration of organs varied with time, while long-time severe drought could cause carbon depletion [45]. It is well known that soluble sugars have functions for osmotic regulation, thus NSC can be used as an osmotic buffer in plants [46]. In addition, plants can convert insoluble starch into soluble sugars, helping to reduce the plant’s osmotic potential to maintain cell expansion during drought [47,48]. Similarly, in this study, reduced starch levels and increased soluble sugar levels in roots were found during drought. However, the drought did not significantly affect soluble sugars in the leaf and stem, which is in line with some previous studies [7,36]. Furthermore, our results showed that SS/NSC values were increased, which is consistent with other reports that plants in drought can maintain a greater proportion of soluble sugars [49]. The increase in root soluble sugars demonstrates that osmotic regulation in roots is likely a strategy for C. camphora to cope with drought [39]. Soluble sugars can also be used for tissue and organ reconstruction under suitable conditions [50], such as restoring hydraulic conductance.
In contrast to drought, NSC in leaf and stem were increased under flooding stress due to the accumulation of starch, which is consistent with some studies [13,42]. Starch accumulation in leaves is a tolerance strategy in Picea mariana and Larix laricina [51]. The increase in starch concentration in leaves and stems may also be due to the decrease in phloem transport and the source–sink effect [17]. Nonetheless, the effect of flooding–drought alternation on NSC was different from drought and flooding alone stress. Under flooding–drought alternation, SS and ST of all organs did not show notable changes compared with the well-watered treatment. However, we found that starch and NSC of leaves and stems were lower but those of roots were higher than flooding stress, which may be related to the restoration of phloem transport from shoots to roots during the recovery phase.
Proline can contribute to osmoregulation and avoidance of damage to the cell membrane [52]. The present study shows that proline concentration was increased in leaves under drought compared with the well-watered treatment at the end of the experiment, suggesting that C. camphora can acclimate to drought stress through osmotic adjustment by proline accumulation, which is consistent with some previous studies [53]. In our study, flooding stress did not have effects on proline concentration, which agrees with a previous study with Populus deltoides [16].
Carbon, nitrogen, and phosphorus are the basic nutrient elements for plants, and most nutrients taken up by roots from the soil depend on the soil's water condition [54]. Moreover, transpiration plays an important contribution in the absorption of nutrients by roots and transport to aboveground parts [54]. Soil drought can inhibit the rate of nutrient diffusion from the soil matrix to the surface of absorbing root and microbial mineralization. The transpiration rate also decreases due to the closure of stomata. Therefore, the nutrients in leaves can be lower [55]. However, in the present research, drought stress did not affect nutrient (C, N, P) concentration in leaves of C. camphora, indicating that the nutrient status was not impacted under the intensity and duration of the drought stress applied in this study. Moreover, flooding stress can affect soil fertility and inhibit plant absorption capacity, thus affecting plant nutrient status. Plenty of results showed that the nutrients of leaves were reduced under flooding conditions [56]. For example, the nitrogen concentration in the leaves of citrus seedlings was decreased. However, nutrients (C, N, P) of leaves were not impacted by flooding stress in the present study, which is also supported by other reports [56,57]. Martínez et al. found no differences in carbon concentration of leaves, twigs, and roots between control and flooding treatments, which may be related to the inhibition of growth and regulation of respiration rates [58]. Additionally, flooding–drought alternation treatment had similar effects on nutrients compared to drought and flooding, suggesting that flooding–drought alternation did not induce the nutrient deficiency of C. camphora.
Generally, our study had certain limitations. This study used potted seedlings in short-term experiments. Thus, the results may differ from experiments with mature trees in the field and results cannot be simply extrapolated to larger scales. Given potentially greater vulnerability to water stress in seedlings compared with mature trees, trait responses could be less pronounced in mature trees. Nevertheless, if the experiment lasts much longer, trait responses could be much more significant. Despite these limitations, this short-term study demonstrated differences in seedling response to different water stress conditions, which help to enhance the physiological understanding of how tree seedlings can cope with multiple environmental stressors.
5. Conclusions
Our findings suggest that drought stimulated root growth while flooding favored leaf growth. Therefore, C. camphora seedlings displayed distinct growth strategies in response to these two contrasting water stressors. The decrease in Asat under both drought and flooding stress was affected by both stomatal and non-stomatal limitations. The regulation of gs improved water use efficiency by promoting leaf water potential declines and the accumulation of abscisic acid (ABA) in leaves, particularly under drought conditions. In addition, drought stress led to some degree of carbon depletion in both stem and root, whereas flooding stress primarily induced carbon depletion in the root. Furthermore, C. camphora could restore physiological responses once stresses (flooding, drought) were relieved. Our results have shown the different effects of drought and flooding on the growth and physiology of C. camphora seedlings, emphasizing the need to consider specific water stress conditions in future studies and providing a theoretical basis for better management of this tree species under variable rainfall patterns. To comprehensively understand the effects of variable water conditions on tree species, further longer-term field studies are warranted, incorporating a broader range of water stress scenarios.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f14071343/s1, Figure S1: Dynamics of Rd of C. camphora seedlings under treatments; Figure S2. SPAD of C. camphora seedlings under treatments on different days.; Figure S3. WUEi as a function of Ψx on different treatments; Figure S4. SS/NSC of leaf (A), stem (B), root (C) and ST/NSC of leaf (D), stem (E), root (F) under treatments.
Author Contributions
H.D. conceived the study, D.W. conducted the data collection, Y.Z. and D.W. analyzed the data. Y.Z. and D.W. wrote the manuscript with input from H.D. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by grants from the Natural Science Talent Funding of Guizhou University (202132).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- IPCC. Climate Change 2021 the Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2021. [Google Scholar]
- Kahraman, A.; Kendon, E.J.; Chan, S.C.; Fowler, H.J. Quasi-Stationary intense rainstorms spread across Europe under climate change. Geophys. Res. Lett. 2021, 48, e2020GL092361. [Google Scholar] [CrossRef]
- Adams, H.D.; Zeppel, M.J.B.; Anderegg, W.R.L.; Hartmann, H.; Landhäusser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.B.; Chen, M.X.; Wang, L.N.; Sasidharan, R.; Voesenek, L.A.; Xiao, S. Multi-stress resilience in plants recovering from submergence. Plant Biotechnol. J. 2022, 21, 466–481. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Chen, J.; Duan, B.; Wang, M.; Korpelainen, H.; Li, C. Intra-and inter-sexual competition of Populus cathayana under different watering regimes. Funct. Ecol. 2014, 28, 124–136. [Google Scholar] [CrossRef]
- Duan, H.L.; Li, Y.; Xu, Y.; Zhou, S.; Liu, J.; David, T.T.; Liu, J. Contrasting drought sensitivity and post-drought resilience among three co-occurring tree species in subtropical China. Agric. For. Meteorol. 2019, 272–273, 55–68. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Santos, D.L.; Moutinho-Pereira, J.M.; Goncalves, B.C.; Ferreira, H.F.; Correia, C.M. Immediate responses and adaptative strategies of three olive cultivars under contrasting water availability regimes: Changes on structure and chemical composition of foliage and oxidative damage. Plant Sci. 2006, 170, 596–605. [Google Scholar] [CrossRef]
- Boughalleb, F.; Hajlaoui, H. Physiological and anatomical changes induced by drought in two olive cultivars (cv Zalmati and Chemlali). Acta Physiol. Plant 2011, 33, 53–65. [Google Scholar] [CrossRef]
- Azeem, A.; Sun, J.; Javed, Q.; Jabran, K.; Du, D. The effect of submergence and eutrophication on the trait’s performance of Wedelia trilobata over its congener native Wedelia chinensis. Water 2020, 12, 934. [Google Scholar] [CrossRef] [Green Version]
- Ferner, E.; Rennenberg, H.; Kreuzwieser, J. Effect of flooding on C metabolism of flood-tolerant (Quercus robur) and non-tolerant (Fagus sylvatica) tree species. Tree Physiol. 2012, 32, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Jia, W.; Ma, M.; Chen, J.; Wu, S. Plant morphological, physiological and anatomical adaption to flooding stress and the underlying molecular mechanisms. Int. J. Mol. Sci. 2021, 22, 1088. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Cheng, R.; Xiao, W.; Guo, Q.; Wang, N. Effect of off-season flooding on growth, photosynthesis, carbohydrate partitioning, and nutrient uptake in Distylium chinense. PLoS ONE 2014, 9, e107636. [Google Scholar] [CrossRef] [PubMed]
- Else, M.A.; Coupland, D.; Dutton, L.; Jackson, M.B. Decreased root hydraulic conductivity reduces leaf water potential, initiates stomatal closure and slows leaf expansion in flooded plants of castor oil (Ricinus communis) despite diminished delivery of ABA from the roots to shoots in xylem sap. Physiol. Plant. 2001, 111, 46–54. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Rennenberg, H. Molecular and physiological responses of trees to waterlogging stress. Plant Cell Environ. 2014, 37, 2245–2259. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.F.; Yang, F. Sex-specific responses to winter flooding, spring waterlogging and post-flooding recovery in Populus deltoides. Sci. Rep. 2017, 7, 2534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, A. Responses to flooding of plant water relations and leaf gas exchange in tropical tolerant trees of a black-water wetland. Front. Plant Sci. 2013, 4, 106. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Sun, H.; Sheng, J.; Jin, S.; Zhou, F.; Hu, Z.; Diao, Y. Transcriptome, physiological and biochemical analysis of Triarrhena sacchariflora in response to flooding stress. BMC Genet. 2019, 20, 88. [Google Scholar] [CrossRef]
- Loreti, E.; Valeri, M.C.; Novi, G.; Perata, P. Gene regulation and survival under hypoxia requires starch availability and metabolism. Plant Physiol. 2018, 176, 1286–1298. [Google Scholar] [CrossRef]
- Irfan, M.; Hayat, S.; Hayat, Q.; Afroz, S.; Ahmad, A. Physiological and biochemical changes in plants under waterlogging. Protoplasma 2010, 241, 3–17. [Google Scholar] [CrossRef]
- Zuniga-Feest, A.; Bustos-Salazar, A.; Alves, F.; Martinez, V.; Smith-Ramirez, C. Physiological and morphological responses to permanent and intermittent waterlogging in seedlings of four evergreen trees of temperate swamp forests. Tree Physiol. 2017, 37, 779–789. [Google Scholar] [CrossRef] [Green Version]
- Guillermo, N.D.; Silvia, E.M.; María, E.R.; Virginia, M.C.L. Physiological responses to alternative flooding and drought stress episodes in two willow (Salix spp.) clones. Can. J. For. Res. 2016, 47, 174–182. [Google Scholar]
- CMA Climate Change Centre. Blue Book on Climate Change in China Science; CMA Climate Change Centre: Beijing, China, 2022. [Google Scholar]
- Li, X.H.; Zhang, Q.; Zhang, D.; Ye, X.C. Investigation of the drought-flood abrupt alternation of streamflow in Poyang Lake catchment during the last 50 years. Hydrol. Res. 2017, 48, 1402–1417. [Google Scholar] [CrossRef]
- Duan, H.; Wang, D.; Wei, X.; Huang, G.; Fan, H.; Zhou, S.; Wan, S. The decoupling between gas exchange and water potential of Cinnamomum camphora seedlings during drought recovery and its relation to ABA accumulation in leaves. J. Plant Ecol. 2020, 13, 683–692. [Google Scholar] [CrossRef]
- Choi, D.S.; Hwang, B.K. Proteomics and functional analyses of pepper abscisic acid–responsive 1 (ABR1), which is involved in cell death and defense signaling. Plant Cell 2011, 23, 823–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Lu, C.; Feng, X.; Guo, G. Effect of autumn foliar application of fertilizers on cold resistance of Red Globe grapes. J. Northwest AF Univ. 2014, 42, 126–132. [Google Scholar]
- Duan, H.L.; Huang, G.; Zhou, S.; Tissue, D.T. Dry mass production, allocation patterns and water use efficiency of two conifers with different water use strategies under elevated [CO2], warming and drought conditions. Eur. J. For. Res. 2018, 5, 605–618. [Google Scholar] [CrossRef]
- Kasurinen, A.; Koikkalainen, K.; Anttonen, M.J.; Possen, B.; Oksanen, E.; Rousi, M.; Holopainen, T. Root morphology, mycorrhizal roots and extramatrical mycelium growth in silver birch (Betula pendula Roth) genotypes exposed to experimental warming and soil moisture manipulations. Plant Soil 2016, 407, 341–353. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C. Individual and interactive responses of woody plants’ biomass and leaf traits to drought and shade. Glob. Ecol. Biogeogr. 2022, 32, 35–48. [Google Scholar] [CrossRef]
- Eziz, A.; Yan, Z.; Tian, D.; Han, W.; Tang, Z.; Fang, J. Drought effect on plant biomass allocation: A meta-analysis. Ecol. Evol. 2017, 7, 11002–11010. [Google Scholar] [CrossRef]
- Yulianti, Y.; Sudrajat, D.J. Morphological responses, sensitivity and tolerance indices of four tropical trees species to drought and waterlogging. 2016, 17, 110–115. Biodivers. J. Biol. Divers.
- Parad, G.A.; Zarafshar, M.; Striker, G.G.; Sattarian, A. Some physiological and morphological responses of Pyrus boissieriana to flooding. Trees 2013, 27, 1387–1393. [Google Scholar] [CrossRef]
- Striker, G.G. Time is on our side: The importance of considering a recovery period when assessing flooding tolerance in plants. Ecol. Res. 2012, 27, 983–987. [Google Scholar] [CrossRef]
- Yan, W.; Zhong, Y.; Shangguan, Z. Elevated temperature exacerbates the effects of drought on the carbon and hydraulic characteristics of Robinia pseudoacacia seedlings. Agric. For. Meteorol. 2020, 280, 107794. [Google Scholar] [CrossRef]
- Creek, D.; Lamarque, L.J.; Torres-Ruiz, J.M.; Parise, C.; Burlett, R.; Tissue, D.T.; Delzon, S. Xylem embolism in leaves does not occur with open stomata: Evidence from direct observations using the optical visualization technique. J. Exp. Bot. 2020, 71, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; Cochard, H. Hydraulic failure defines the recovery and point of death in water-stressed conifers. Plant Physiol. 2009, 149, 575–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juzoń, K.; Czyczyło-Mysza, I.; Ostrowska, A.; Marcińska, I.; Skrzypek, E. Chlorophyll fluorescence for prediction of yellow lupin (Lupinus luteus L.) and pea (Pisum sativum L.) susceptibility to drought. Photosynthetica 2019, 57, 950–959. [Google Scholar] [CrossRef] [Green Version]
- Estill, K.; Delaney, R.H.; Smith, W.K.; Ditterline, R.L. Water relations and productivity of alfalfa leaf chlorophyll variants. Crop Sci. 1991, 31, 1229–1233. [Google Scholar] [CrossRef]
- Yang, Y.J.; Bi, M.H.; Nie, Z.F.; Jiang, H.; Liu, X.D.; Fang, X.W.; Brodribb, T.J. Evolution of stomatal closure to optimize water-use efficiency in response to dehydration in ferns and seed plants. New Phytol. 2021, 230, 2001–2010. [Google Scholar] [CrossRef]
- Rengifo, E.; Tezara, W.; Herrera, A. Water relations, chlorophyll a fluorescence, and contents of saccharides in tree species of a tropical forest in response to flood. Photosynthetica 2005, 43, 203–210. [Google Scholar] [CrossRef]
- Martorell, S.; Diaz-Espejo, A.; Medrano, H.; Ball, M.C.; Choat, B. Rapid hydraulic recovery in Eucalyptus pauciflora after drought: Linkages between stem hydraulics and leaf gas exchange. Plant Cell Environ. 2014, 37, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, H.; Trumbore, S. Understanding the roles of non-structural carbohydrates in forest trees from what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Liu, H.; Qi, Y.; Liu, F.; Zhu, X. Patterns in nonstructural carbohydrate contents at the tree organ level in response to drought duration. Glob. Chang. Biol. 2020, 26, 3627–3638. [Google Scholar] [CrossRef]
- Thalmann, M.; Santelia, D. Starch as a determinant of plant fitness under abiotic stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.S.; Gear, L.; Hultine, K.R.; Koch, G.W.; Ogle, K. Non-structural carbohydrate dynamics associated with antecedent stem water potential and air temperature in a dominant desert shrub. Plant Cell Environ. 2020, 43, 1467–1483. [Google Scholar] [CrossRef]
- Sapes, G.; Demaree, P.; Lekberg, Y.; Sala, A. Plant carbohydrate depletion impairs water relations and spreads via ectomycorrhizal networks. New Phytol. 2021, 229, 3172–3183. [Google Scholar] [CrossRef]
- Blumstein, M.; Gersony, J.; Martinez-Vilalta, J.; Sala, A. Global variation in nonstructural carbohydrate stores in response to climate. Glob. Chang. Biol. 2022, 29, 1854–1869. [Google Scholar] [CrossRef]
- Galvez, D.A.; Landhausser, S.M.; Tyree, M.T. Root carbon reserve dynamics in aspen seedlings: Does simulated drought induce reserve limitation? Tree Physiol. 2011, 31, 250–257. [Google Scholar] [CrossRef]
- Islam, M.A.; Macdonald, S. Ecophysiological adaptations of black spruce (Picea mariana) and tamarack (Larix laricina) seedlings to flooding. Trees 2004, 18, 35–42. [Google Scholar] [CrossRef]
- Ahmed, C.B.; Rouina, B.B.; Sensoy, S.; Boukhris, M.; Abdallah, F.B. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2009, 67, 345–352. [Google Scholar] [CrossRef]
- Ditmarová, L.; Kurjak, D.; Palmroth, S.; Kmet, J.; Střelcová, K. Physiological responses of Norway spruce (Picea abies) seedlings to drought stress. Tree Physiol. 2010, 30, 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, E.C.; Nogueira, R.J.M.C.; Da Silva, M.A.; de Albuquerque, M.B. Drought stress and plant nutrition. Plant Stress 2011, 5, 32–41. [Google Scholar]
- Gessler, A.; Schaub, M.; McDowell, N.G. The role of nutrients in drought-induced tree mortality and recovery. New Phytol. 2017, 214, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Alcántara, B.; Jover, S.; Quinones, A.; Forner-Giner, M.Á.; Rodríguez-Gamir, J.; Legaz, F.; Iglesias, D.J. Flooding affects uptake and distribution of carbon and nitrogen in citrus seedlings. J. Plant Physiol. 2012, 169, 1150–1157. [Google Scholar] [CrossRef]
- Wang, A.F.; Roitto, M.; Lehto, T.; Sutinen, S.; Heinonen, J.; Zhang, G.; Repo, T. Photosynthesis, nutrient accumulation and growth of two Betula species exposed to waterlogging in late dormancy and in the early growing season. Tree Physiol. 2017, 37, 767–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey-Serres, J.; Voesenek, L. Flooding stress: Acclimations and genetic diversity. Annu. Rev. Plant Biol. 2008, 59, 313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Schematic diagram of experimental design. WW seedlings were watered to field capacity throughout the experiment, while DS seedlings were desiccated by withholding water completely. Pots of FS seedlings were placed inside larger plastic buckets by maintaining the water level 5 cm above the soil surface. FDF treatments consisted of three stages: first, seedlings were treated with flooding (same with FS) until stomatal conductance (gs) declined to near 0 mol m−2 s−1 (day 11); seedlings were then moved out from the buckets and left desiccated by withholding water, in order to create the condition for post-flooding recovery followed by drought stress; finally, when gs declined to 0 mol m−2 s−1 due to the drought stress (day 36), potted seedlings were treated with another round of flooding towards the end of the experiment (day 40).
Figure 1.
Schematic diagram of experimental design. WW seedlings were watered to field capacity throughout the experiment, while DS seedlings were desiccated by withholding water completely. Pots of FS seedlings were placed inside larger plastic buckets by maintaining the water level 5 cm above the soil surface. FDF treatments consisted of three stages: first, seedlings were treated with flooding (same with FS) until stomatal conductance (gs) declined to near 0 mol m−2 s−1 (day 11); seedlings were then moved out from the buckets and left desiccated by withholding water, in order to create the condition for post-flooding recovery followed by drought stress; finally, when gs declined to 0 mol m−2 s−1 due to the drought stress (day 36), potted seedlings were treated with another round of flooding towards the end of the experiment (day 40).

Figure 2.
Dynamics of Ψpd (A), Ψm (B), Ψx (C) of C. camphora seedlings under different water treatments. Values are means ± SE (n = 3–4). Treatments: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation). Asterisks indicate significant difference between WW and other water stress treatments (p < 0.05).
Figure 2.
Dynamics of Ψpd (A), Ψm (B), Ψx (C) of C. camphora seedlings under different water treatments. Values are means ± SE (n = 3–4). Treatments: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation). Asterisks indicate significant difference between WW and other water stress treatments (p < 0.05).

Figure 3.
Dynamics of Asat (A), E (B), gs (C), and WUEi (D) of C. camphora seedlings under different water treatments. Values are means ± SE (n = 3–4). Asterisks indicate significant difference between WW and other water stress treatments (p < 0.05). Treatments: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation).
Figure 3.
Dynamics of Asat (A), E (B), gs (C), and WUEi (D) of C. camphora seedlings under different water treatments. Values are means ± SE (n = 3–4). Asterisks indicate significant difference between WW and other water stress treatments (p < 0.05). Treatments: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation).

Figure 4.
gs as a function of ABA (A) and Asat as a function of SPAD (B) under different treatments. Treatments: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation).
Figure 4.
gs as a function of ABA (A) and Asat as a function of SPAD (B) under different treatments. Treatments: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation).

Figure 5.
Asat (A), gs (B), E (C) as a function of Ψm under different treatments. Curve fitting was only performed for the DS treatment. Treatments: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation).
Figure 5.
Asat (A), gs (B), E (C) as a function of Ψm under different treatments. Curve fitting was only performed for the DS treatment. Treatments: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation).

Figure 6.
SS, ST, and NSC of leaf (A–C), stem (D–F), and root (G–I) under different treatments. Values are means ± SE (n = 3–4). Different letters indicate significant difference between different treatments (p < 0.05). Treatments: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation).
Figure 6.
SS, ST, and NSC of leaf (A–C), stem (D–F), and root (G–I) under different treatments. Values are means ± SE (n = 3–4). Different letters indicate significant difference between different treatments (p < 0.05). Treatments: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation).

Figure 7.
Dynamics of ABA (A) and proline (B) of C. camphora seedlings under different treatments. Values are means ± SE (n = 3–4). NA means no data available. Different letters indicate significant difference between different treatments (p < 0.05). Treatments: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation).
Figure 7.
Dynamics of ABA (A) and proline (B) of C. camphora seedlings under different treatments. Values are means ± SE (n = 3–4). NA means no data available. Different letters indicate significant difference between different treatments (p < 0.05). Treatments: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The total biomass and biomass allocation of C. camphora seedlings at the end of the experiment. Values are means ± SE (n = 4–5). Treatment: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation). Different letters in the same row denote significant differences among treatments (p < 0.05).
Table 1.
The total biomass and biomass allocation of C. camphora seedlings at the end of the experiment. Values are means ± SE (n = 4–5). Treatment: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation). Different letters in the same row denote significant differences among treatments (p < 0.05).
| Parameter | Treatments | |||
|---|---|---|---|---|
| WW | DS | FS | FDF | |
| Total biomass (g) | 40.83 ± 0.60 a | 23.58 ± 0.57 b | 30.30 ± 1.75 b | 23.88 ± 4.05 b |
| Root-to-shoot ratio | 0.55 ± 0.02 b | 0.68 ± 0.06 a | 0.41 ± 0.04 c | 0.63 ± 0.02 ab |
| Root mass ratio | 0.35 ± 0.01 a | 0.40 ± 0.02 a | 0.29 ± 0.02 b | 0.39 ± 0.01 a |
| Stem mass ratio | 0.39 ± 0.01 ab | 0.42 ± 0.03 ab | 0.36 ± 0.02 b | 0.46 ± 0.03 a |
| Leaf mass ratio | 0.26 ± 0.01 b | 0.18 ± 0.02 c | 0.35 ± 0.03 a | 0.15 ± 0.03 c |
Table 2.
The leaf nutrient concentration and carbon isotopic composition of C. camphora seedlings at the end of the experiment. Values are means ± SE (n = 3–4). Treatment: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation). Different letters in the same row denote significant differences among the treatments (p < 0.05).
Table 2.
The leaf nutrient concentration and carbon isotopic composition of C. camphora seedlings at the end of the experiment. Values are means ± SE (n = 3–4). Treatment: WW (well-watered), DS (drought stress), FS (flooding stress), FDF (flooding–drought alternation). Different letters in the same row denote significant differences among the treatments (p < 0.05).
| Parameter | Treatments | |||
|---|---|---|---|---|
| WW | DS | FS | FDF | |
| C concentration (mg/g) | 280.06 ± 40.99 a | 285.81 ± 2.96 a | 272.23 ± 8.58 a | 282.52 ± 7.16 a |
| N concentration (mg/g) | 9.68 ± 0.30 a | 10.73 ± 0.71 a | 9.02 ± 1.09 a | 8.99 ± 0.66 a |
| P concentration (mg/g) | 1.99 ± 0.12 a | 1.99 ± 0.03 a | 2.01 ± 0.05 a | 2.17 ± 0.14 a |
| C:N (%) | 28.75 ± 3.34 a | 26.85 ± 1.61 a | 31.21 ± 4.30 a | 31.68 ± 1.76 a |
| C:P (%) | 141.60 ± 21.44 a | 143.41 ± 0.77 a | 135.75 ± 6.55 a | 131.41 ± 9.37 a |
| N:P (%) | 4.89 ± 0.33 a | 5.38 ± 0.37 a | 4.48 ± 0.46 a | 4.16 ± 0.26 a |
| 13C (‰) | −34.27 ± 0.40 b | −32.98 ± 0.40 a | −33.68 ± 0.33 ab | −33.52 ± 0.25 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, Y.; Wang, D.; Duan, H. Effects of Drought and Flooding on Growth and Physiology of Cinnamomum camphora Seedlings. Forests 2023, 14, 1343. https://doi.org/10.3390/f14071343
AMA Style
Zhao Y, Wang D, Duan H. Effects of Drought and Flooding on Growth and Physiology of Cinnamomum camphora Seedlings. Forests. 2023; 14(7):1343. https://doi.org/10.3390/f14071343
Chicago/Turabian StyleZhao, Yongju, Defu Wang, and Honglang Duan. 2023. "Effects of Drought and Flooding on Growth and Physiology of Cinnamomum camphora Seedlings" Forests 14, no. 7: 1343. https://doi.org/10.3390/f14071343
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.