In this study, we morphologically and phylogenetically analyze and report 22 Ganoderma species from the GMS, namely, G. adspersum, G. applanatum, G. australe, G. calidophilum, G. ellipsoideum, G. flexipes, G. gibbosum, G. heohnelianum, G. hochiminhense, G. leucocontextum, G. lucidum, G. multiplicatum, G. multipileum, G. myanmarense, G. orbiforme, G. philippii, G. resinaceum, G. sichuanense, G. sinense, G. subresinosum, G. williamsianum, and G. tsugae. Of these 22 species, 12 were from Yunnan Province, China; three were from Laos; three species, two new records, and one new species were from Myanmar; 15 species and four new records were from Thailand; and one new species was from Vietnam. The phylogenetic and morphological analyses results of the 22 Ganoderma species are detailed below.
3.2. Taxonomy
Ganoderma P. Karst., Revue Mycologique Toulouse. 3(9): 17 (1881)
= Dendrophagus Murrill, Bull. Torrey bot. Club. 32(9): 473 (1905)
= Elfvingia P. Karst., Bidr. Känn. Finl. Nat. Folk. 48: 333 (1889)
= Friesia Lázaro Ibiza, Revista Real Acad. Ci. Madrid. 14: 587 (1916)
= Ganoderma subgen. Trachyderma Imazeki, Bull. Tokyo Sci. Mus. 1: 49 (1939)
= Tomophagus Murrill, Torreya. 5: 197 (1905)
= Trachyderma (Imazeki) Imazeki, Bull. Gov. Forest Exp. Stn Tokyo. 57: 97 (1952)
Type species: Ganoderma lucidum (Leyss: Fr.) Karst.
Notes: (≡) is homotypic, or nomenclatural, synonyms, (=) is heterotypic, or taxonomic, synonyms.
Description: Basidiomes annual, dimidiate, sessile or sub-stipitate to stipitate. Pileus subdimidiate to dimidiate, flabelliform, perennial, stipitate or sessile. Pileus surface non-laccate (dull) or weakly to strongly laccate, glossy, shiny, smooth, spathulate, shallow, furrows, sulcate, several layers thick, with thin- to thick-cuticle cells or cuticle of clavate end cells, thicker at the base than the margin, thin- to thick-crust overlaying the pileus, consistency hard, consistency hard, light weight when dried. Pileus color variable, light yellow to yellow, light brown, slightly brown to dark brown, sometimes homogeneous reddish gray to reddish-yellow. Context brown to dark brown, grayish orange to orange, sometimes grayish-yellow, mostly soft, sometimes spongy to firm-fibrous. Hymenophore di-trimitic, heterogeneous, non-septate or septate, usually yellow, slightly light orange, or light brown to brown, sometimes with melanoid bands. Tubes are hard, woody when dried. Tube layers single or stratified, pale to purplish-brown, almost hyaline with clamp connections, occasionally branched at apex, thin- to thick-walled. Stipe central or lateral, glossy with a distinct cuticle. Margin actively growing, entirely white when fresh, round, soft and smooth when young, slippery when touched from youth to maturity, and tough when broken. Pores 4–7 in number per mm, angular, entire, subcircular to circular, regular, cream or white when young, light yellow, light orange to brown when mature. Pore surface usually white to cream when fresh, turning yellowish-white to pale yellow on drying, some sections reddish gray to brown, and brownish gray when wet.
Hyphal structure: Hyphal system di-trimitic, including generative, skeletal, and binding hyphae; mostly generative hyphae with clamp connections, hyaline, brown, non-septate, or septate, often with long and tapering branches. Basidia broadly ellipsoid, tapering abruptly at the base. Cystidia absent. Basidiospores broadly to narrowly ellipsoid or oblong, sometimes globose to subglobose, with double walls, truncate apex, apical germ pore present, usually with light brown to brown endosporium, hyaline exosporium with thin inter-walled pillars, hyaline endosporium with thick outer walls, and some very thin exosporium.
Ecology: mostly on hardwoods, trunks, and stumps, occurring on several different living tree host species.
Notes: Justo et al. [
103] treated Ganodermataceae as a synonym of Polyporaceae and included the genus
Ganoderma under Polyporaceae. Later, Cui et al. [
104] excluded
Ganoderma from Polyporaceae due to the presence of double-walled basidiospores, unlike Polyporaceae. So, the distinctiveness of the genus
Ganoderma lies in the presence of double-walled and truncate basidiospores. Species with a laccate, glossy surface are present in both Ganodermataceae and Polyporaceae as centrally and laterally stipitate species.
3.2.1. Taxonomy of Ganoderma from China
Ganoderma angustisporum J.H. Xing, B.K. Cui and Y.C. Dai, Mycokeys 34: 98 (2018)
Taxonomy and phylogenic analysis were described in Xing et al. [
37]
Notes: Ganoderma angustisporum is characterized by annual, sessile, broadly basidiomes, strongly laccate on the upper surface of basidiomes, white pore surfaces, and almond-shaped, slightly truncate, narrow 9.0–11.3 × 4.0–5.2 µm basidiospores. It is a group of white-rot fungi that predominantly grow on living Casuarina equisetifolia in Fujian Province, China.
Ganoderma applanatum (Pers.) Pat., Hymenomyc. Eur. (Paris): 143 (1887) (
Figure 4)
≡ Boletus lipsiensis Batsch, Elenchus fungorum. Continuatio prima.: 183, t. 25:130 (1786)
≡ Scindalma lipsiense (Batsch) Kuntze, Revisio generum plantarum. 3(2): 518 (1898)
≡ Polyporus lipsiensis (Batsch) E.H.L. Krause, Basidiomycetes Rostochienses.: 54 (1928)
≡ Agaricus lipsiensis (Batsch) E.H.L. Krause, Basidiomycetum Rostochiensium, Suppl. 4: 142 (1932)
= Boletus applanatus Pers., Observationes mycologicae. 2: 2 (1799)
= Polyporus merismoides Corda, Deutschlands Flora, Abt. III. Die Pilze Deutschlands. 3: 139 (1837)
= Polyporus stevenii Lév., Annls Sci. nat., Bot.: 91 (1844)
= Polyporus leucophaeus Mont., Sylloge generum specierumque plantarum cryptogamarum.: 157 (1856)
= Polyporus leucophaeum Mont. (1856)
= Polyporus incrassatus Berk., Journal of the Linnean Society. Botany. 16: 41 (1877)
= Polyporus concentricus Cooke, Grevillea. 9(49): 13 (1880)
= Fomes gelsicola Berl., Malpighia. 3: 373 (1889)
= Fomes nigriporus Lázaro Ibiza, Revista de la Real Academia de Ciencias Exactas Fisicas y Naturales Madri. 14: 662 (1916)
= Ungularia subganodermica Lázaro Ibiza, Revista de la Real Academia de Ciencias Exactas Fisicas y Naturales Madri. 14: 674 (1916)
= Fomes longoporus Lloyd, Mycological Writings. 6(62): 940 (1920)
Facesoffungi number: FoF 06249
Description: Basidiomes annual, perennial, sessile. Pileus 1.5–5.8 cm in length, 0.5–4.5 cm in width, and up to 1.5 cm thick at the base, sessile (without stipe), perennial, subdimidiate, sub-flabelliform to flabelliform, usually flat, convex, imbricate, umbonate or uneven, rarely ungulate, glabrous when present, broadly attached when mature, often with undefined concentric zones at the center that extend to the margin, thick at the base, slightly soft at the margin when mature. Pileus surface shiny, silky, smooth, and soft when young, hard when old, frequently furrowed and shallow sulcate, undulating, somewhat spathulate to uneven on the upper surface when mature, covered by irregularly ruptured thick crust, slightly non-laccate (dull) and faded from when mature to old, compact and hard when mature, woody to corky when old. Pileus color is usually homogenous with grayish-orange (6B3–6B5) at the center, slight brownish-orange (6C4), orange white (6A2), to pale orange (6A3), with yellowish-gray (4B2) at the margin when mature. Context up to 0.3–1 cm thick at the base, mostly light brown (7D5), brown (6E8) to dark brown (7F6–7F8) of cuticle cells, with walls varying in thickness to subsolid hyphae, some fibrous pithy context, usually separated by layers of context tissue at the base, and some occurred woody lines. Tube woody, hard, often dark brown (7F7–7F8) when dried, with sulcate at different levels. Stipe almost sessile and broadly attached when present, with a differentiated zone at the point of attachment. Margin up to 1 cm thick, white (5A1), yellowish-gray (4B2) when mature, turns light brown (6D4) to brown (6E8) when scratched or bruised, often slippery when wet, soft when young, thinner than the center. Pore 4–6 in number per mm, subcircular to circular, sometimes angular. Pore surface initially white (7A1), grayish-orange (7C3–7C4) when mature, turning to light brown (7D6) to brown (7D8) when scratched or bruised.
Hyphal structure: Hyphal system trimitic; generative hyphae 0.8–2.6 µm ( = 2.1, n = 30) in diam, almost hyaline, with clamp connections, abundant, thin-walled and occasionally thick-walled; skeletal hyphae 2.1–4.6 µm width (n = 30), usually thick-walled, hyaline, sometimes branched; binding hyphal 1.6–3.3 µm width (n = 30), thick-walled and occasionally thin-walled, branched, and intertwined with the skeletal hyphae. Pileipellis a hymeniderm, grayish brown (6E4), which is composed of apically acanthus-like branched cells. Basidiospores mostly ellipsoid with double walls, with a size range of (9.8–)10.4–11.1–11.9(–12.1) × (7.3–)8.0–8.6–9.2(–9.9) μm, ( = 11.3 × 8.7 μm, n = 50) μm, with Q = 1.79–1.86, L = 11.23 µm, W = 6.12 µm (including myxosporium), (6.2–)7.6–8.6–9.7(–10.4) × (5.0–)5.8–7.1–8.2(–8.9) μm ( = 8.6 × 7.1 μm, n = 50) μm, with Q = 1.19–1.24, L = 8.59 µm, W = 7.12 µm (excluding outer myxosporium), brownish-orange (7D4) to brown (7D7–7D8) in KOH, and reddish-brown (8E6) to dark brown (8F4) in Melzer’s reagent. Basidia 14–20 × 8–10 μm, with four sterigmata.
Ecology: Solitary on stump of Machilus yunnanensis.
Specimens examined: CHINA, Yunnan Province, Baoshan, 25°09′35″ N, 99°09′49″ E, 1973 m elev., 11 November 2017, T. Luangharn, HKAS 107254, MFLU 19-2188.
Notes: Ganoderma lipsiense has been treated by some researchers as the correct name for
G. applanatum [
15].
G. applanatum (=
G. lipsiense) belongs in the subgenus
Elfvingia, which is characterized by distinctive non-laccate species, a thin and acute margin of the pileus, and unbranched terminal endings of skeletal hyphae, with ellipsoid basidiospores [
47,
105,
106,
107].
G. applanatum causes white butt rot on angiosperm trees and is widely distributed in China [
64]. Hence, our specimen of
G. applanatum, collected from a temperate region of China, is described based on morphological characteristics and molecular phylogenetic data. Our results agree well with those of Ryvarden and Gilbertson [
105].
Ganoderma australe (Fr.) Pat., Bull. Soc. mycol. Fr. 5(2–3): 65 (1889) (
Figure 5)
≡ Polyporus australis Fr., Elenchus Fungorum. 1: 108 (1828)
≡ Fomes australis (Fr.) Cooke, Grevillea. 14(69): 18 (1885)
≡ Placodes australis (Fr.) Quél., Enchiridion Fungorum in Europa media et praesertim in Gallia Vigentium. 171 (1886)
≡ Fomes applanatus var. australeis (Fr.) Cleland and Cheel, Journal of Proceedings of the Royal Society of New South Wales. 51: 518 (1918)
≡ Ganoderma applanatum subsp. australe (Fr.) Bourdot and Galzin, Bulletin de la Société Mycologique de France. 41: 184 (1925)
≡ Ganoderma applanatum f. australe (Fr.) Bourdot and Galzin, Bulletin de la Société Mycologique de France. 41: 184 (1925)
≡ Elfvingia australis (Fr.) G. Cunn., Bulletin of the New Zealand Department of Industrial Research. 164: 256 (1965)
= Polyporus tornatus Pers., Botanique (Nagpur). 5: 173 (1827)
= Polyporus scansilis Berk., Journal of the Linnean Society. Botany. 16: 53 (1877)
= Fomes annularis Lloyd, Mycol. Writ. 4(40): 6 (1912)
= Ganoderma tornatum var. tornatum (Pers.) Bres., Hedwigia. 53(1–2): 55 (1912)
= Fomes konigsbergii Lloyd (1915)
= Fomes polyzonus Lloyd, Synopsis of the genus Fomes. (7): 269 (1915)
= Fomes pseudoaustraleis Lloyd, Synopsis of the genus Fomes. (7): 269 (1915)
= Fomes undatus Lázaro Ibiza, Revista de la Real Academia de Ciencias Exactas Fisicasy Naturales Madri. 14: 661 (1916)
Facesoffungi number: FoF 06242
Description: Basidiomes annual, perennial, subdimidiate, sessile. Pileus 14–28 cm in length, 12–32 cm in width, and 1.4–3.2 cm thick. Pileus flabelliform, spathulate, subdimidiate, umbonate, single, sulcate, large, obtuse from host, radial from the center extending the margin, broadly attached, often thick at the center, slightly soft at the margin, consistency hard, and tough to break when dried. Pileus surface convex, corky, furrowed, spathulate, mostly umbonate or uneven, non-laccate (dull) on maturity or in old, usually slippery where the new hyphae are in active development (margin), slightly concentrically sulcate at the center toward margin, smooth, covered with thick and hard crust, irregularly ruptured crust overlying the surface, woody, and corky when dried, with cracked crust when mature, and tough to break when dried. Pileus color often brown (6E7–6E8) at the base, reddish-orange (7B7–7B8), brownish-orange (7C6–7C8), almost covered with grayish-red (8C4–8C5) on the upper surface when old, slight reddish-brown (8F8, 9E7–9E8) close to the margin. Context up to 0.5–2 cm thick near stipe, fibrous, composed of coarse loose fibrils, brown (6D7–6D8) to dark brown (6F7), with reddish-brown (8D8–8D9) coarse loose fibrils, covered with thick crust. Tube 0.4–1.5 cm long, brown (7D8) to dark brown (6F8). Stipe sessile, broadly attached. Margin soft when young, slippery when fresh, blunt when mature, white (4A1). Pore 4–6 in number per mm, subcircular to circular, sometimes angular. Pore surface initially white (4A1), slight to pale yellow (4A3) when mature, turned brownish-orange (6C7–6C8) when scratched or bruised or discolored when touched.
Hyphal structure: Hyphal system trimitic, dense and hard, thick-walled, typically with narrow lumen, flexuous, and many branches, usually brownish-orange (6C5–6C7) in KOH; generative hyphae 2.2–3.8 µm broad (n = 30), thin-walled, hyaline, tapering at branch, with clamp connections; skeletal hyphae 2.9–4.2 µm broad (n = 30), sometimes branched, nearly solid, thick-walled; binding hyphae 2.6–4.0 µm broad (n = 30), thick-walled, branched, more or less solid; hymenial with sword-like apices in the context. Pileipellis a hymeniderm, usually brownish-orange (6C5) to brown (6E8), composed of apically acanthus-like branched cells. Basidiospores mostly ellipsoid to broadly ellipsoid, double walls, (6.2-)7.1–9.4–10.4(-11.8) × (5.2-)6.0–7.4–8.9(-9.7) μm ( = 9.4 × 7.4 μm, n = 50) μm, with Q = 1.24–1.30, L = 9.42 µm, W = 7.43 µm (including myxosporium), (5.3-)6.7–7.8–9.6(-10.5) × (4.5-)5.1–5.7–6.3(-7.2) μm ( = 7.8 × 5.7 μm, n = 50) μm, with Q = 1.31–1.38, L = 7.85 µm, W = 5.83 µm (excluding outer myxosporium), overlaid by hyaline, apically brown, bearing a fine, distinct, short, echinulae truncate, turgid vesicular appendix, inner wall orange (6B8) to brownish-orange (6C8) or light brown (7D5–7D6) to brown (7E7–7E8), outer wall mostly reddish-brown (8E7–8E8, 8F7) in 5% KOH.
Habitat: Solitary, growing on living Neocinnamomum delavayi (Lec.) H. Liou. tree or, decaying stump, and living Fagus spp.
Specimens examined: CHINA, Yunnan Province, Kunming Botanical Garden, 25°08′39″ N, 102°44′30′ E, 1956 m, 27 September 2016, T. Luangharn, HKAS 97397.
Notes: Ganoderma australe belongs under the subgenus
Elfvingia [
38]. This fungus was initially described from the Pacific Islands [
108].
G. australe belongs to the
G. applanatum-
australe complex [
2]. Ganoderma australe was established as a non-laccate (dull) pilei. The type specimen of this fungus is missing, while the neotype specimen is available in Europe [
105].
Ganoderma applanatum and
G. australe from Europe have been confused based on the macro-characteristic features [
107]. The typification of
G. australe remains unresolved and was exemplified by several authors [
61,
109,
110,
111], and its similar cultural characteristics also showed the phenotypic plasticity in morphological level and a higher level of nucleotide divergence in the ITS rDNA region that made
G. australe a complex species [
45].
Ganoderma australe is distinguished from
G. applanatum by the larger dimensions of its basidiospores, different stipe features, thickness of the cuticle, and color of the context layer, all of which were considered in delimiting
G. applanatum and
G. australe [
2,
105,
107].
G. applanatum is confined to northern temperate regions, while
G.
australe can be found in tropical and subtropical regions [
112,
113]. There are reports on the occurrence in Australia [
14], China [
30], New Zealand [
114], southern India [
51,
95], Taiwan, PRC [
115], and Thailand [
30,
59].
Ganoderma australe is a cosmopolitan species, which is known to cause white rot on woody material. It shows parasitic or pathogenic behavior on a wide range of both dead and living broadleaved deciduous trees [
116,
117,
118].
Ganoderma australe is distributed worldwide, especially in tropical regions [
14].
Ganoderma calidophilum J.D. Zhao, L.W. Hsu and X.Q. Zhang, Acta Mycologica Sinica 19: 270 (1979) (
Figure 6)
Facesoffungi number: FoF 06244
Description: Basidiomes annual, stipitate, subdimidiate. Pileus 3–7 cm in length, 2–4 cm in width, and 0.2–1 cm thick. Pileus subdimidiate to dimidiate, spathulate, stipitate, sulcate, umbonate, radial from the center extending to the margin, tough to break when dried, often thick at the center, slightly soft at the margin, and light in weight when dried. Pileus surface corky, convex, furrowed, glabrous, glossy, incised, shiny, spathulate, shallow, sulcate when fresh, umbonate or uneven, laccate and glossy when mature, weakly laccate when old and in regions of developing hyphae (margin), slightly concentrically sulcate, layers smooth at the center when young, irregularly ruptured crust overlying the context, and tough to break when dried. Pileus color usually homogenous with brownish-red (8C7–8C8), brownish-red (9C7–9C8), reddish-brown (9D6–9D7) center, extending brownish-orange (6C7–6C8) toward the stipe, brownish-red (9C8) from the center to light brownish-orange (6C8), and usually light brown (6D8) at the margin when old. Context up to 0.2–0.6 cm thick near stipe, dry, fibrous, composed of coarse loose fibrils, brownish-orange (6C5–6C8) upper layers when fresh, brown (6D7) at lower layers, dark brown (8F7) when dried, covered with thin crust, trimitic hyphal system. Tube 0.3–0.9 cm in length, brown (7D8). Stipe 5–14 cm long, cylindrical, almost stipitate with broadly, irregularly ruptured crust overlying, strongly laccate with brown (7D8) when mature, dark brown (8F8) when old, and woody or corky when dried. Margin soft when young, laccate when mature, weakly laccate to laccate when old, blunt when old, usually light brown (6D8) when mature to old. Pore 4–5 in number per mm, subcircular to circular, sometimes angular. Pore surface initially pale orange (5A3) to brownish-orange (6D8) when mature, discolored when touched, brown (6E8) when scratched or bruised.
Hyphal structure: Hyphal system trimitic hyphal, usually brownish-orange (6C5–6C7) in KOH; generative hyphae 1.2–3.2 µm broad (n = 30), thin-walled, hyaline, without clamp connections; skeletal hyphae 3.2–6.4 µm broad (n = 30), sometimes branched, nearly solid, thick-walled, without clamp connections; binding hyphae 2.4–5.2 µm broad (n = 30), usually thin to thick-walled, many branches, nearly solid, hymenial with sword-like apices in the context. Basidiospores mostly ellipsoid to broadly ellipsoid, with double wall, with a size range of (7.4-)8.5–11.9–12.6(-13.7) × (6.3-)7.2–8.3–9.1(-9.6) μm ( = 11.9 × 8.3 μm, n = 50) μm, with Q = 1.39–1.45, L = 11.92 µm, W = 8.35 µm (including myxosporium), (6.8-)7.6–10.4–11.3(-12.8) × (5.4-)6.3–7.0–7.6(-8.1) μm ( = 10.4 × 7.1 μm, n = 50) μm, with Q = 1.43–1.49, L = 10.39 µm, W = 7.12 µm (excluding outer myxosporium), overlaid by hyaline, apically and echinulae, truncate, turgid vesicular appendix, inner wall orange (6B8) to deep orange (6A8), reddish-orange (7A8, 7B7–7B8) or yellowish-red (8B8), outer wall usually reddish-brown (8D7–8D8; 8E8) in 5% KOH.
Habitat: Solitary, near the hardwood root of Castanopsis spp., living tree of Machilus yunnanensis.
Specimens examined: CHINA, Yunnan Province, Baoshan, 25°06′29″ N, 99°08′29″ E, 1973 m elev., 11 November 2017, T. Luangharn, MFLU 19-2174.
Notes: Ganoderma calidophilum is a species originally described from Hainan Province, China, by Zhao [
119]. Several reports have confirmed that this fungus is mentioned in Hainan Province polypore diversity checklists [
13,
61,
120,
121]. This fungus is distinctive in these forms, featuring a laccate pileus with broadly ellipsoid basidiospores with double walls, and it is widely found across subtropical and tropical Asia [
108,
119,
122]. Wang and Wu [
112] suggested that
G. calidophilum is a synonym of
G. flexipes. However, the evaluated
G. calidophilum and
G. flexipes are different in terms of pileus color, pileus shape and size, context, and basidiospore size [
112]. In this study, we present our
G. calidophilum collection from Yunnan Province, China, based on taxonomic and phylogenetic analyses. Our strain is similar to the described strain of Wang and Wu [
119], Zhao et al. [
119], and Bi et al. [
123].
Ganoderma flexipes Pat., Bulletin de la Société Mycologique de France. 23(1): 75 (1907) (
Figure 7)
≡ Fomes flexipes (Pat.) Sacc. and Traverso, Sylloge Fungorum. 19: 710 (1910)
≡ Polyporus flexipes (Pat.) Lloyd, Synopsis of the stipitate Polyporoids. (7): 104 (1912)
Facesoffungi number: FoF 06245
Description: Basidiomes annual or perennial, stipitate. Pileus 0.5–3.2 cm in length, 0.5–3 cm broad, up to 0.5 cm thick at the base. Pileus stipitate, sub-reniform to reniform, or subflabellate to flabellate, concentrically sulcate zones with tuberculate, glabrous when young to maturity, bumps when mature, often tough to break when dried. Pileus surface shiny, smooth, and soft when young, frequently furrowed and shallow sulcate on upper surface, undulating, somewhat spathulate to uneven when mature, covered by an irregularly ruptured thin crust, faded or weakly laccate when young, and laccate when mature., and woody when old. Pileus color usually homogenous with reddish-brown (8E8) to dark brown (9F7–9F8) at the center, slight to the margin from mature to old. Context up to 0.1–0.6 cm thick at the base, very dry, brown (7D7–7D8) to reddish-brown (8E7), containing fibrous pithy context and corky when old. Tube hard, often dark brown (7F8). Stipe almost 3–12 cm in length, 0.3–1.5 in width, sub-cylindrical to cylindrical, often dark brown (7F8), and strongly laccate from mature to old. Margin soft when young, laccate when mature, some wavy, often light brown (6D5–6D6) on the upper surface. Pore 4–5 in number per mm, subcircular to circular, sometimes angular. Pore surface initially grayish-orange (6B4–6B6), turns brown (7D7) to reddish-brown (8D5–8D7) when scratched or bruised, discolored when touched.
Hyphal structure: Hyphal system trimitic, bearing clamp connections, hyaline, thick-walled, tapering branch, some swollen differentiated zone at the point of attachment; generative hyphae (1.8-)2.2–2.9–3.4(-3.8) μm broad (n = 30), thin-walled, hyaline, unbranched, with clamp connections; skeletal hyphae (3.0-)3.8–4.8–5.4(-6.2) μm broad (n = 30), with walls varying in thickness, with subsolid, binding hyphae (2.2-)2.8–3.8–4.5(-5.1) μm broad (n = 30), usually thick-walled, appearing alongside Bovista hyphae, and many branches, usually light yellow (4A4–4A5) to yellowish-orange (4B8) of thin-walled and orange (6A7) to deep orange (6A8) of thick-walled in Melzer’s reagent. Pileipellis a hymeniderm, dark brown (6F8), composed of apically acanthus-like branched cells. Basidiospores mostly ellipsoid to broadly ellipsoid with double wall at maturity, (8.1-)8.8–9.7–10.6(-11.2) × (6.1-)6.6–7.7–9.7(-10.4) μm ( = 9.7 × 7.7 μm, n = 50) μm, with Q = 1.08–1.15, L = 9.68 µm, W = 7.72 µm (including myxosporium), (7.3-)7.8–8.3–8.7(-9.2) × (4.0-)4.6–5.4–5.8(-6.2) μm ( = 10.2 × 6.4 μm, n = 50) μm, with Q = 1.51–1.57, L = 8.34 µm, W = 5.39 µm (excluding outer myxosporium), overlaid by hyaline, dextrinoid, echinulae, inner wall echinulate brown (5D8, 7E6–7E8), and outer wall usually dark brown (7E8) to reddish-brown (8E6–8E8) in Melzer’s reagent.
Habitat: Solitary on the decaying hardwood of Pinus spp.
Specimens examined: CHINA, Yunnan Province, Baoshan, 25°06′29″ N, 99°08′29″ E, 1973 m elev., November 2017, T. Luangharn, MFLU 19-2189.
Notes: Ganoderma flexipes is originally described from Vietnam by Patouillard [
124]. It has been recorded from China, India, Laos, Nepal, and Pakistan [
4,
13,
30,
110,
122].
G. flexipes is characterized by its small reddish-brown pileus, long and thin stipe, usually reddish-brown to dark brown context, and ellipsoid or ovoid basidiospores [
125]. Among the Chinese
Ganoderma species,
G. flexipes is one of the most similar species to
G. sichuanense as they share a reddish-brown pileal surface, similar basidiospores, and cuticle cells [
4]. Our
G. flexipes from China is very similar to the description of Ryvarden [
125] and Hapuarachchi et al. [
30], and basidiospores are within the range of 9.7–10.2 × 6.4–7.7 μm.
Ganoderma gibbosum (Blume and T. Nees) Pat., Ann. Jard. Bot. Buitenzorg, suppl. 1: 114 (1897) (
Figure 8)
≡ Polyporus gibbosus (Blume and T. Nees)., Nov. Act. Academiae Caesareae Leopoldino Carolinae Germanicae Naturae Curiosorum. 13: 19, t. 4(1–4) (1826)
≡ Fomes amboinensis var. gibbosus (Blume and T. Nees) Cooke, Grevillea. 13(68): 118 (1885)
≡ Fomes gibbosus (Blume and T. Nees) Sacc. Syll. Fung. 6: 156 (1888)
≡ Scindalma gibbosum (Blume and T. Nees) Kuntze., Revisio generum plantarum 3(2): 518 (1898)
Facesoffungi number: FoF 06246
Description: Basidiomes annual or perennial, sessile. Pileus 8–21 cm in length, 6–13 cm in width, and 1–3.5 cm thick, convex, imbricate, umbonate, uneven, ungulate, subflabellate, subdimidiate, usually round when present, primordial, somewhat round and plump when young, somewhat imbricate, when seen from above flabelliform (fan-shaped), broadly attached, thick at the base, slightly soft at the margin when mature. Pileus surface non-laccate (dull), furrowed, incised, sulcate, smooth when young, usually silky, soft, and slippery surface when fresh, undulating on the upper surface, somewhat spathulate to uneven, with a crust (0.2–0.4 mm), woody from mature to older, and lined or cracked crust occurs when old. Pileus color usually homogenous with grayish-orange (6B3–6B6), brownish-orange (6C5–6C6), and brown (6D7–6D8) at the base extending to the margin of mature fruiting bodies. Context up to 0.5–1.8 cm thick, compact and hard, trimitic hyphal, with clamp connections, hyaline, with walls varying in thickness with simple septa, composed of narrow and sparingly branched; generative hyphae 1.3–3.2 µm broad (n = 30) with hyaline; skeletal hyphae 2.8–4.7 µm broad (n = 30), usually thick-walled; binding hyphal 2.1–4.2 µm width (n = 30) with walls varying in thickness. Hymenophore reddish-brown (8D7). Tube layers 0.4–1.2 cm in length. Stipe almost sessile and broadly attached when present. Margin blunt-edged, wavy, slippery from young, softer, and often white (8A1) when youth to maturity, and light brown (6D5) when old, the present yellow line between the edge of the margin and close to the underside of basidiomes. Pore 4–7 in number per mm, subcircular to circular. Pore surface white (11A1) when present, pale yellow (4A3) to grayish-yellow (4B3–4B4) when scratched or bruised, discolored when touched.
Hyphal structure: Hyphal system trimitic hyphal, with clamp connections, usually reddish-brown (8D7–8D8); generative hyphae (1.2-)1.5–2.4–3.0(-3.6) μm broad (n = 30), thin-walled and hyaline; skeletal hyphae (2.7-)3.2–3.6–4.2(-4.8) μm broad (n = 30), dextrinoid, abundant thick wall; binding hyphae (2.5-)3.0–3.5–4.0(-4.4) μm broad (n = 30), thick wall, branched, usually intertwined the generative and skeletal hyphae, mostly dark brown near the tube layers, appearing alongside Bovista-type ligative hyphae, hymenial, sword-like apices at the context. Pileipellis a hymeniderm, dark brown (6D8), composed of apically acanthus-like branched cells, dextrinoid. Basidiospores mostly ellipsoid to broadly ellipsoid or oblong with double walls, (5.8-)6.2–7.2–8.4(-9.2) × (5.4-)5.7–5.4–6.8(-7.7) μm ( = 7.3 × 5.6 μm, n = 50) μm, with Q = 1.48–1.52, L = 7.32 µm, W = 5.68 µm (including myxosporium), (4.8-)5.2–6.0–6.7(-7.2) × (4.6-)4.9–5.5–5.7(-6.2) μm ( = 6.2 × 5.6 μm, n = 50) μm, with Q = 1.08–1.14, L = 6.24 µm, W = 5.67 µm (including myxosporium), overlaid by hyaline, dextrinoid, echinulae, echinulate brown inner wall, light yellow (4A4–4A5) to grayish-yellow (4B5–4B6) in 5% KOH. Basidia not seen.
Habitat: Solitary on decaying hardwood of Machilus yunnanensis, living tree of Albizia mollis and Pinus spp.
Specimens examined: CHINA, Yunnan Province, Kunming Institute of Botany garden, 25°08′39″ N, 102°44′30″ E, 1956 m elev., 31 December 2016, T. Luangharn, HKAS 97411.
Notes: Ganoderma gibbosum belongs to the family Ganodermataceae, which was first described in Australia [
126].
G. gibbosum has been recorded from China [
40], India [
95], Korea [
92], Laos [
30], and Thailand [
30]. This species is distinctive in having non-laccate basidiomes and ellipsoids with double-walled basidiospores [
40].
Ganoderma gibbosum has been reported to cause white rot and several other diseases in hard woods [
7] and is widely distributed in both tropical and temperate areas [
4]. It was considered to be a subspecies of
G. applanatum [
4], while
G. applanatum was the earlier name of
G. australe [
127].
Ganoderma australe and
G. gibbosum were renamed as
G. incrassatum based on their monophyletic origin [
128] since it had been well recognized that
G. applanatum was synonymized with
G. applanatum.
Ganoderma leucocontextum T.H. Li, W.Q. Deng, Sheng H. Wu, D.M. Wang and H.P. Hu, Mycotaxon 56: 82 (2015) (
Figure 9)
Facesoffungi number: FoF 06247
Description: Basidiomes flabelliform, subdimidiate, stipitate. Pileus 6–14 cm in length, 4–12 cm in width, and 1–3.2 cm thick. Pileus flabelliform, spathulate, stipitate, subdimidiate to dimidiate, umbonate, somewhat semicircular, plump, concentrically sulcate zone, broad and thick at the base, mostly radial from the center extending to the margin, tough to break when dried, often thick at the center, slightly soft at the margin, light in weight when dried, and not woody. Pileus surface convex, furrowed, imbricate, incised, glossy, shiny, spathulate, shallow sulcate when fresh, umbonate or uneven, usually smooth layers at center when young to age, non-laccate to weakly laccate when present, strongly laccate and glossy when mature, weakly laccate where the new hyphae are in active development (margin), irregularly ruptured crust overlying the context, and tough to break when dried. Pileus color usually homogenous with orange (6A7) and deep orange (6A8) at the center toward stipe, extending deep orange (5A8) from the center, slight deep yellow (4A8) where the new hyphae are in active development when mature, usually red (11B7–11B8) at the center, and orange (6A7) to deep orange (6A8)–(6B8) extending to the upper margin surface from mature to old. Context up to 0.3–2.4 cm thick near stipe, white context when fresh, yellowish-white (1A2) when dried, soft and fibrous, trimitic hyphal, with clamp connections, hyaline, with walls varying in thickness with simple septa, and unbranched. Tubes 0.3–1.2 cm in length. Stipe 3–10 cm in length, 4–7 cm in width, sub-cylindrical to cylindrical, almost stipitate, broad at the base, some presented short stipitate, strongly laccate with dark brown (8F7–8F8) to grayish ruby (12E6–12E7) when mature, and grayish brown (8E4) when old. Margin wavy, softer, slippery when young, laccate when mature, strongly laccate when old, orange yellow (4A7) to deep yellow (4D8) where the new hyphae are in active development, deep orange (6A7–6A8) to brown (6D8) from mature to old. Pore 4–6 in number per mm, subcircular, some circular, or angular. Pore surface white (11A1) when present, yellowish-white (2A2) when mature, brownish-orange (6C7–6C8) when scratched or bruised, discolored when touched.
Hyphal structure: Hyphal system trimitic, usually golden brown (5D7), yellowish-brown (5D8) to reddish-brown (8D7–8D8) in KOH; generative hyphae 2.3–5.2 µm broad (n = 30), thin-walled, hyaline, with clamp connections; skeletal hyphae 2.6–5.5 µm broad (n = 30), thick-walled, unbranched or rearly branched; binding hyphae 1.8–4.2 µm width (n = 30), usually thin to thick-walled, branched, hymenial with sword-like apices in the context. Basidiospores mostly ellipsoid to broadly ellipsoid, double walls, (8.8-)9.3–10.7–11.4(-12.6) × (6.8-)7.4–8.3–8.7(-9.2) μm ( = 10.5 × 8.4 μm, n = 50) μm, with Q = 1.22–1.28, L = 10.52 µm, W = 8.41 µm (including myxosporium), (7.8-)8.1–8.5–8.8(-9.1) × (5.2-)5.7–6.0–6.5(-6.9) μm ( = 8.3 × 6.2 μm, n = 50) μm, with Q = 1.32–1.38, L = 8.34 µm, W = 6.18 µm (excluding outer myxosporium), overlaid by hyaline, apically echinulae, truncate, some turgid, vesicular appendix, inner walled echinulate, golden yellow (5B7), grayish-orange (5D6) to yellowish-brown (5D7), outer walled reddish-brown in 5% KOH. Cystidia absent. Cultures characteristics white mycelial after incubation at 30 °C for 10 days.
Habitat: Solitary, on the decaying hardwood of unknown tree.
Specimens examined: CHINA, Yunnan Province, Baoshan, 25°09′35″ N, 99°09′49″ E, 1973 m elev., 26 October 2016, J. Xu, HKAS 97401.
Notes: Ganoderma leucocontextum was introduced by Li et al. [
62] from the Tibet Autonomous Region of China. This species can be easily recognized by its stipitate, white context, thick stipe, broadly ellipsoid basidiospores (9–12.5 × 7–9 μm), coarse echinulae, mostly regular cuticle hyphae, and its deciduous wood habitat [
62]. The holotype is similar to
G. lucidum from Europe [
62]; however, the illustrated differences in macro-characteristics of the European
G. lucidum are smaller basidiospores (7–12 × 6–8 μm), a deeper-colored context that is usually rust-colored, becoming dark purple to brown in older portions [
105]. Additionally,
G. leucocontextum also resembles the widely cultivated
G. lucidum (
G. lingzhi) in East Asia [
62], but the Chinese
G. lucidum has a deeper-colored context and is even darker near the tube layer, with shorter cutis elements (20–40 × 7–15 μm) and smaller spores (8–11.5 × 5.5–8.5 μm) (including myxosporium) than
G. leucocontextum leucocontextum [
5]. Our
G. leucocontextum collection from Hainan Province agrees well with the descriptions provided by Li et al. [
62].
Ganoderma lucidum (Curtis) P. Karst., Revue Mycologique Toulouse. 3(9): 17 (1881) (
Figure 10)
≡ Boletus rugosus Jacq., Flora Austriaca. 2: 44, f. 169 (1774)
≡ Boletus lucidus Curtis, Fl. Londinensis. 4: 72, t. 224 (1781)
≡ Polyporus lucidus (Curtis) Fr., Systema Mycologicum. 1: 353 (1821)
≡ Grifola lucida (Curtis) Gray, A natural arrangement of British plants. 1: 644 (1821)
≡ Fomes lucidus (Curtis) Cooke, Grevillea. 13(68): 118 (1885)
≡ Placodes lucidus (Curtis) Quél., Enchiridion Fungorum in Europa media et praesertim in Gallia Vigentium.: 170 (1886)
≡ Phaeoporus lucidus (Curtis) J. Schröt., Kryptogamen-Flora von Schlesien. 3-1(4): 491 (1888)
= Boletus flabelliformis Leyss., Flora halensis.: 219 (1761)
= Agaricus pseudoboletus Jacq., Miscellanea austriaca ad botanicum, chemiam et historiam naturalem spectantia. 1: 26, t. 41 (1773)
= Boletus obliquatus Bull., Herbier de la France. 1: t. 7 (1781)
= Boletus vernicosus Bergeret, Phytonomatotechnie universelle. 1: 99 (1783)
= Agaricus lignosus Lam., Encyclopédie Méthodique, Botanique. 1-1: 51 (1783)
= Boletus dimidiatus Thunb., Fl. Japonica.: 348, f. 39 (1784)
= Boletus castaneus Weber, Suppl. Fl. hols.: 13 (1787)
= Boletus laccatus Timm, Flora megapolitanae Prodomus exhibeus plantas ductatus Megapolitano.: 269 (1788)
= Boletus crustatus J.J. Planer, Index Plantarum quas in Agro Erfurtensi sponte provenientes.: 280 (1788)
= Agarico igniarium trulla Paulet, Traité des champignons. 2: 95, pl. 10:1-2 (1793)
= Boletus verniceus Brot., Flora Lusitanica. 2: 468 (1804)
= Ganoderma ostreatum Lázaro Ibiza, Revta R. Acad. Cienc. exact. fis. nat. Madr.: 110(1916)
= Ganoderma nitens Lázaro Ibiza, Revta R. Acad. Cienc. exact. fis. nat. Madr.: 104 (1916)
Facesoffungi number: FoF 06250
Description: Basidiomes imbricate, reniform, stipitate. Pileus up to 2–5 cm in length, 2–4 cm in width, and 0.8–2.2 cm thick. Pileus stipitate, reniform, imbricate, irregular, some laterally, and flabelliform with a contracted, concentrically sulcate zone, irregularly ruptured crust overlying the context, radial or branched from the center extending to the margin, tough to break when dried, often thick at the center, slightly soft at margin, and leathery when aged, tough to break when dried. Pileus surface weakly laccate when present, strongly laccate and glossy when mature, weakly laccate where the new hyphae are in active development (margin), smooth layer at the center from young to age, usually furrowed, incised, undulate to sulcate, somewhat spathulate to uneven, some woody or corky when old. Pileus color usually yellowish-red (8B7–8B8) at the center, slight to reddish-orange (7B7–7B8), and orange (6A7–6A8) on upper pileus surface. Context up to 0.4–1.4 cm thick at the base, abundant thick-walled, subsolid hyphae, concentric lines of various shade, bearing clamp connections, light brown (6D6) to brown (6D8, 6E8), presenting dark brown (6F8) melanoid bands. Tube hard, often brown (7D7) to dark brown (7F7). Stipe up to 8–16 cm in length, up to 0.6–1.8 cm in width, central stipe, cylindrical, thick with uneven at the base (up to 1.8 cm), usually dark brown (7F7–7F8), laccate, and cracked when old. Margin often 0.4–1 cm, orange (6A7–6A8) on upper surface, and reddish-yellow (4A8) under surface, thin and soft than the center. Pore (75–)110–145(–165) μm, circular, some angular, 4–6 in number per mm. Pore surface white (11A1) to light brown (7D6), turning brown (7D7–7D8) to dark brown (6F6) when scratched or bruised.
Hyphal structure: Hyphal system trimitic, with clamp connections, hyaline, thin-walled with abundant thick-walled with simple septa, sparingly branched, swollen by melanoid bands, usually pale orange (5A3), light orange (5A5), to reddish-orange (8A7–8A8) in KOH; generative hyphae up to 1.7–3.2 μm broad (n = 30), almost hyaline, usually thin to thick-walled, with clamp connections, and sparingly branched and flexuous; skeletal hyphae 3.0–6.4 μm broad (n = 30), usually thick-walled with clamp, and abundantly branched and flexuous; binding hyphae 2.0–5.6 μm broad (n = 30), usually thick-walled with abundant branches, and occurring melanoid bands. Basidiospores ellipsoid to broadly ellipsoid, some globose with double walls, with a truncate apex, with double wall, mostly overlaid by hyaline myxosporium, eusporium bearing fine, short, and distinct, coarse, echinulae, hyaline, turgid, vesicular appendix, (7.7-)8.4–9.4–10.6(-11.5) × (5.2-)5.9–6.3–7.1(-8.4) μm, ( = 9.5 × 6.4 μm, n = 50) μm, with Q = 1.47–1.52, L = 9.52 µm, W = 6.34 µm (including myxosporium), (6.0-)6.9–7.3–8.1(-8.5) × (4.6-)4.9–5.3–5.8(-6.2) μm ( = 7.5 × 5.2 μm, n = 50) μm, with Q = 1.41–1.47, L = 7.52 µm, W = 5.24 µm (excluding outer myxosporium), brownish-orange (6C8), (6D8) to brown (6E5) of endosporium (inner wall) with brown (7E7–7E8) exosporium (outer wall) in Congo red, brownish-orange (6C8) in 5% KOH, and yellowish-brown in Melzer’s reagent.
Habitat: Solitary, on decaying hardwood of Quercus sp. in the native forest.
Specimens examined: CHINA, Yunnan Province, Honghe, 23°21′50″ N, 103°22′24″ E, 874 m elev., 15 August 2017, T. Luangharn, MFLU 19-2161.
Notes: Ganoderma lucidum (Curtis) P. Karst. was originally reported from temperate England [
2]. Previously, it was characterized as
Boletus lucidus Curtis and then
Polyporus lucidus (Curtis) Fr. (1821) [
1]. The species
P. lucidus was characterized by having a laccate pileus and stipe. The molecular phylogenetic analyses indicated that the
G. lucidum from Europe is not conspecific to the Chinese
G. lucidum; thus, the European
G. lucidum remained the true
G. lucidum, and the Chinese
G. lucidum was proposed as
G. lingzhi [
4], and most of the collections named
G. lucidum in East Asia were not conspecific with the
G. lucidum found in Europe [
129].
Ganoderma lucidum is relatively common in Europe, while its geographic distribution in East Asia, East Africa, Europe, North America, Asia, and other parts of the world is largely unknown [
38].
Several studies of
Ganoderma have used the name
G. lucidum for any laccate
Ganoderma species, as
Ganoderma are ly highly variable, often resulting in taxonomic and phylogenetic confusion, especially with regards to
G. lucidum [
38]. The taxonomy of
Ganoderma has been a constant topic of debate due to the high levels of phenotypic plasticity in its species. Several characteristics of
Ganoderma are similar to
G. lucidum, such as
G. multipileum [
39],
G. oregonense [
42],
G. resinaceum [
41],
G. tsugae,
G. lucidum,
G. sichuanense, and
G. sinense [
4,
32,
33,
35,
40,
130] from China. Cao et al. [
4] have clarified a different new species, Chinese
G. lucidum as
G. lingzhi, which has an East Asian distribution. The most striking characteristics that differentiate
G. lucidum from
G. lingzhi are the presence of melanoid bands in the context, a yellow pore surface, and thick dissepiments (80–120 μm) at maturity [
4]. The molecular evidence reveals
G. lucidum and
G. sinense as two clear different species [
130].
In China, the first-reported
G. lucidum was illustrated based on collections from Guizhou Province [
124]. Then, Teng [
131] reported more collections from different regions of China, and many subsequent collections have been reported [
4,
13,
47,
121,
132]. Recently, this fungus has been reported to be distributed worldwide based on gross similarity of features, e.g., in Europe [
105], Asia [
13,
133], America [
134,
135], and Africa [
112]. Our collection from Yunnan Province, China, also agrees well with the descriptions provided from Asia.
Ganoderma multiplicatum (Mont.) Pat., Bulletin de la Société Mycologique de France 5: 74 (1889) (
Figure 11)
≡ Polyporus multiplicatus Mont., Annales des Sciences Naturelles Botanique. 1: 128 (1854)
≡ Fomes multiplicatus (Mont.) Cooke, Grevillea. 14 (69): 18 (1885)
≡ Scindalma multiplicatum (Mont.) Kuntze, Revisio generum plantarum. 3 (2): 519 (1898)
Facesoffungi number: FoF 06251
Description: Basidiomes annual or perennial, stipitate with short base. Pileus 1.5–7.5 cm in length, 0.5–4 cm in width, and up to 1.5 cm thick at the base. Pileus dimidiate, flabelliform, reniform, usually flat, convex, imbricate, umbonate or uneven, rarely ungulate, glabrous when present, often with undefined concentric zones at the center that extend to the margin, and thick at the base, slightly soft at margin when mature. Pileus surface shiny, silky, smooth, and soft when young, non-laccate (dull) when mature, hard and woody when old, frequently furrowed and shallow sulcate, undulating, somewhat spathulate to uneven on upper surface when mature, covered by irregularly ruptured thick crust, slightly dull and faded when mature to old, compact and hard when mature, woody to corky from mature to old. Pileus color usually homogenous with grayish-orange (6B3) at the center slight to brownish-orange (6C4) and pale orange (6A3), usually yellowish-gray (4B2) at the margin when mature, and brown (6E8) when dried. Context up to 0.4–1 cm thick at the base, mostly brown (6E8) to dark brown (7F6–7F8) of cuticle cells, and dark brown (6F6) melanoid bands, thick-walled, some fibrous pithy context, usually separated by layers of context tissue at the base. Tube woody hard, often with dark brown (7F7–7F8) when dried, with sulcate at different levels. Stipe short stipitate, dark brown (7F7), and a differentiated zone at the point of attachment. Margin up to 1 cm thick, initially white (5A1), yellowish-gray (4B2) when mature, turns light brown (6D4) to brown (6E8) when scratched or bruised, often slippery when wet, softer when young, thinner than the center. Pore 4–7 in number per mm, subcircular to circular, some angular. Pore surface initially white (7A1) to yellowish-white (1A2), becoming pale orange (5A3) when mature, light brown (7D6) to brown (7D8) when handled, scratched, and bruised.
Hyphal structure: Hyphal system trimitic; generative hyphae 2.1–4.8 µm ( = 2.2, n = 30) in diam, clamp, almost hyaline, thin to thick-walled, composed of narrow and spare branches; skeletal hyphae 3.2–6.5 µm width (n = 30), usually thick-walled, hyaline, some branched and intertwined hyphae; binding hyphal 2.4–5.7 µm width (n = 30), thick-walled, many branches, and comprised Bovista-type ligative hyphae. Pileipellis a hymeniderm, brown (6E8), composed of apically acanthus-like branched cells, dextrinoid. Basidiospores mostly ellipsoid with double walls, (7.8-)8.7–10.8–12.2(-13.3) × (6.9-)7.4–9.1–10.0(-10.7) μm, ( = 10.7 × 9.1 μm, n = 50) μm, with Q = 1.15–1.22, L = 10.79 µm, W = 9.13 µm (including myxosporium), (5.4-)5.9–6.6–7.1(-7.7) × (4.9-)5.4–5.8–6.2(-6.7) μm ( = 6.6 × 5.8 μm, n = 50) μm, with Q = 1.11–1.17, L = 6.64 µm, W = 5.82 µm (excluding outer myxosporium), inner walled deep orange (6A8), brownish-orange (7C7–7C8) in KOH, outer walled dark brown (7E6), dark brown (7E8) to reddish-brown (8E8) in KOH. Cutis usually composed of clavate cells.
Habitat: Solitary on stump of Quercus spp.
Specimens examined: CHINA, Yunnan Province, Jinning District, 24°41′17″ N, 102°13′15″ E, 1912 m elev., 8 October 2017, T. Luangharn, MFLU 19-2152.
Notes: Ganoderma multiplicatum was originally collected from French Guyana [
2]. This species has a distinctive form with its reddish-black pileus, a not fully homogenous context, tuberculate hyphal ends in cuticle cells, with small subglobose to broadly ellipsoid basidiospores (7–8 × 5–6 μm) [
7,
136,
137,
138].
Ganoderma multiplicatum has been considered most similar to
G. chalceum [
113], also considered a synonym of
G. subamboinense Henn. [
136], but Correia de Lima et al. [
139] illustrated that
G. chalceum and
G. subamboinense are in different clades, suggesting they are not synonymous. Our
G. multiplicatum specimen was collected from Yunnan Province, China. It is similar to the original description, showing ellipsoid basidiospores, while sub-globous basidiospores could not be observed. This species has been reported from Africa [
140], Asia [
61], China [
13,
61,
119,
122], India [
141], Myanmar [
30], Taiwan, PRC [
122], and neotropical regions of Brazil, Colombia, and Venezuela [
138].
Ganoderma resinaceum Boud., Bulletin de la Société Mycologique de France. 5; 72 (1889) (
Figure 12)
≡ Fomes resinaceus (Boud.) Sacc., Sylloge Fungorum. 9: 179 (1891)
≡ Scindalma resinaceum (Boud.) Kuntze, Revisio generum plantarum. 3(2); 519 (1898)
≡ Friesia resinacea (Boud.) Lázaro Ibiza, Revta R. Acad. Cienc. exact. fis. nat. Madr.: 591 (1916)
≡ Ganoderma lucidum subsp. resinaceum (Boud.) Bourdot and Galzin, Bulletin de la Sociètè Mycologique de France. 41; 177 (1925)
≡ Ganoderma lucidum var. resinaceum (Boud.) Maire, Fungi Catalaunici: Contributions á lètude de la Flore Mycologique de la Catalogne: 38 (1933)
= Ganoderma chaffangeonii Pat., Bulletin de la Société Mycologique de France. 5: 74 (1889)
= Polyporus polychromus Copel., Annales Mycologici. 2 (6): 507 (1904)
= Ganoderma praelongum Murrill, North American Flora. 9 (2): 121 (1908)
= Ganoderma argillaceum Murrill, North American Flora. 9 (2): 122 (1908)
= Ganoderma pulverulentum Murrill, North American Flora. 9 (2): 121 (1908)
= Ganoderma subperforatum G.F. Atk., Botanical Gazette Crawfordsville. 46 (5): 337 (1908)
= Ganoderma areolatum Murrill, Bulletin of the New York Botanical Garden. 8: 149 (1912)
= Mensularia vernicosa Lázaro Ibiza, Revista de la Real Academia de Ciencias Exactas Fisicas y Naturales Madri. 14: 740 (1916)
= Ganoderma subtuberculosum Murrill, Lloydia. 7 (4): 326 (1945)
Facesoffungi number: FoF 06252
Description: Basidiomes annual, perennial, short stipitate. Pileus 1.5–12.5 cm in length, 1–7 cm in width, and up to 2 cm thick at the base. Pileus dimidiate, flabelliform, reniform, convex, imbricate, umbonate or uneven, some ungulate, concentric zones at the center that extend to the margin, broadly attached, thick at the base, slightly soft at the margin when mature. Pileus surface glossy, shiny, silky, smooth, and soft when young, laccate when mature, furrowed and shallow sulcate, undulating, somewhat spathulate to uneven on upper surface when mature, covered by irregularly ruptured thin crust, slightly dull and faded when mature to old, compact and hard when mature, woody to corky when mature to old. Pileus color reddish-brown (10E7–10E8) at the center, slight to yellowish-red (8B7–8B8), reddish-orange (7A7–7A8), and light orange (5A5–5A6) closed to the margin, and white (4A1) at the margin. Context up to 0.4–1 cm thick at the base, mostly grayish-yellow (4C6) to dark brown (7F6–7F8) cuticle cells, and dark brown (6F6) melanoid bands, thick-walled, some fibrous pithy context, usually separated by layers of context tissue at the base. Tube woody hard, often dark brown (7F7–7F8) when dried, concolorous with pore surface, and sulcate at different levels. Stipe short stipitate, usually reddish-brown (10E7–10E8), and a differentiated zone at the point of attachment. Margin up to 1.5 cm thick, initially white (5A1), yellowish-gray (4B2) when mature, turning light brown (6D4) to brown (6E8) when scratched or bruised, often slippery when wet, softer when young, thinner than the center. Pore 4–7 in number per mm, angular to circular. Pore surface initially white (7A1) to yellowish-white (1A2), becoming light orange (5A5) when mature, light brown (7D6) to brown (7D8) when handled, scratched, or bruised.
Hyphal structure: Hyphal system trimitic; generative hyphae 2.1–4.7 µm ( = 3.6, n = 30) in diam, clamp, almost hyaline, thin-walled, composed of sparse branches; skeletal hyphae 3.2–6.2 µm width (n = 30), usually thick-walled, hyaline, some branched and intertwined hyphae; binding hyphal 2.8–5.1 µm width (n = 30), thick-walled and occasionally thick-walled, without septate hyphae, many branches, and composed of Bovista-type ligative hyphae. Basidiospores mostly ellipsoid with double walls, (7.6-)8.4–9.4–10.5(-11.3) × (6.5-)7.1–8.4–9.0(-9.8) μm, ( = 9.3 × 8.2 μm, n = 50) μm, with Q = 1.10–1.16, L = 9.31 µm, W = 8.24 µm (including myxosporium), (6.5-)7.1–8.2–9.1(-9.8) × (4.8-)5.3–5.7–6.8(-7.3) μm ( = 8.1 × 5.6 μm, n = 50) μm, with Q = 1.42–1.48, L = 8.13 µm, W = 5.62 µm (excluding outer myxosporium), inner walled orange (5A6) to deep orange (5A7–5A8, 6A8) in KOH and grayish brown (5C5–5C6) in Melzer’s reagent, outer walled dark brown (7E6–7E8) to reddish-brown (8E8) in KOH and light brown (6D5–6D6) to brown (6D7–6D8) in Melzer’s reagent.
Habitat: Solitary, on living tree of Albizia mollis (Wall.) Boiv.
Specimens examined: CHINA, Yunnan Province, Kunming Institute of Botany, 25°08′39″ N, 102°44′30″ E, 1962 m, 12 July 2017, T. Luangharn, MFLU 19-2153.
Notes: Ganoderma resinaceum was introduced by Boudier in 1889 from France [
41]. This species has also been described by Steyaert [
110] and Ryvarden and Gilbertson [
105].
Ganoderma resinaceum is distinctively characterized by variable pileus coloration, a fibrous spongy homogeneous context, larger basidiospores, and an amyloid pileipellis [
7]. This species is considered to have characteristics similar to
G. pfeifferi in its upper crust resinous layers. However, this species has a dark brown to umber context and wider spores. In addition,
G. resinaceum also shares similarities with
G. lucidum, while
G. lucidum possesses a varying light context without a dark zone above the tubes and no resinous layer on the crust [
105,
140].
Ganoderma resinaceum was evaluated to the species complex base on molecular evidence [
38], but in the phylogenetic analysis, it cannot be distinguished from
G. lucidum [
142]. However, several researchers suggested that
G. resinaceum differs from
G. lucidum [
4,
76,
143].
Ganoderma sanduense Hapuar., T.C. Wen and K.D. Hyde, Mycosphere 10, 274 (2019)
Taxonomy and phylogenetic analyses are shown in Hapuarachchi et al. [
30].
Notes: Ganoderma sanduense is characterized by its ferruginous laccate pileus, orbicular, strongly laccate, several layers thick, basidiospores 12.1–13.8 × 9.2–10.5 μm, relatively large broadly ellipsoid to ellipsoid basidiospores, with a light brown eusporium bearing fine, hyaline, short, and distinct echinulae. This fungus is solitary on rotten wood in dry dipterocarp forests and in upper-mixed deciduous forests from Guizhou Province, China.
Ganoderma sichuanense J.D. Zhao and X.Q. Zhang, Acta mycol. sin.: 159 (1983) (
Figure 13)
Facesoffungi number: FoF 06248
Description: Basidiomes annual or perennial, stipitate. Pileus 0.5–3.2 cm in length, 0.5–3 cm in width, up to 1 cm thick at the base. Pileus reniform to circular, or subflabellate when seen from above, concentrically sulcate zones with turberculate, glabrous when youth to maturity, bumps when mature, often tough to break when dried, often with undefined concentric zones at the center that extend to the margin, thick at the center, slightly soft at the margin. Pileus surface shiny, silky, smooth, and soft when young, hard and woody old, frequently furrowed and shallow sulcate on upper surface, undulating, somewhat spathulate to uneven when mature, covered by irregularly ruptured thin crust, and strongly laccate from mature to old. Pileus color usually homogenous with yellowish-red (8A7–8A8) at the center, slight reddish-orange (7A7–7A8), and reddish-brown (8E8) at the deep-sulcate margin from mature to old. Context up to 0.2–0.8 cm thick at the base, some thin-walled, with abundant thick-walled to subsolid hyphae, containing fibrous pithy context, bearing clamp connections, with dark brown (7F7) melanoid bands occurring. Tube hard and woody, thin-walled, frequently branched, with clamped connection, and often dark brown (7F7–7F8) when dried. Stipe up to 3–812 cm in length, up to 0.3–1 cm in width, centrally stipitate, almost sub-cylindrical to cylindrical, concolorous with the pileus, often reddish-brown (8E7–8E8), and strongly laccate from mature to old. Margin soft when young, strongly laccate when mature, some wavy, slippery when wet, smooth, softer, thinner than the base, and soft than the center, often deep orange (5A8) to golden yellow (5B7–5B8) from mature to old. Pore 4–6 in number per mm, subcircular to circular, sometimes angular. Pore surface initially white (11A1), pale yellow (3A3) to yellow (3A7) when mature, turns light brown (7D5), brown (7D7–7D8) to dark brown (7F6–7F8) when scratched or bruised, becoming discolored when touched.
Hyphal structure: Hyphal system trimitic, with clamp connections, hyaline, thin to thick-walled, tapering at branch, sometimes swollen at the attachment point, composed of some narrow hyphae; generative hyphae (1.3–)1.8–2.3–2.6(–2.8) μm broad (n = 30), hyaline, and thin-walled; skeletal hyphae (2.1–)2.5–3.9–4.8(–5.2) μm broad (n = 30) abundant with walls varying in thickness, unbranched, sometimes subsolid; binding hyphae (1.7–)2.1–2.8–3.6(–4.3) μm broad (n = 30), usually with walls varying in thickness, narrow to subsolid, usually presenting as orange white (6A2), pale orange (6A3) to light orange (5A5) of thin-walled, and pale red (6A3) of thick-walled, with subsolid in KOH. Pileipellis a hymeniderm, light brown (6D6), clavate-like cells, dextrinoid. Basidiospores mostly ellipsoid, some oblong with double walls, (8.0-)8.6–9.6–10.5(-11.0) × (6.2-)6.7–8.4–9.6(-10.1) μm ( = 9.5 × 8.3 μm, n = 50) μm, Q = 1.11–1.17, L = 9.49 µm, W = 8.31 µm (including myxosporium), (7.0-)7.4–7.9–8.5(-9.0) × (4.2-)4.5–5.6–5.9(-6.4) μm ( = 7.8 × 5.6 μm, n = 50) μm, with Q = 1.36–1.41, L = 7.80 µm, W = 5.62 µm (excluding outer myxosporium), overlaid by hyaline, dextrinoid, echinulae, inner wall echinulate with grayish-orange (6B5–6B6) to brownish-orange (7D4–7D5), and outer walled usually dark brown (7E8–7E8) to reddish-brown (8E6–8E8) in KOH.
Habitat: Solitary on the living tree of Graucoides schotky.
Specimens examined: CHINA, Yunnan Province, Xishan Forest Park, 24°57′53″ N, 102°53′10″ E, 2013 m elev., 29 October 2016, T. Luangharn, HKAS 97398.
Notes: Ganoderma sichuanense was originally described from the Sichuan Province, China, in 1983 [
5]. However,
G. sichuanense was published in 1983 [
40] but has not been widely used.
Ganoderma sichuanense was verified as “
G. lucidum” (Lingzhi) based on both morphological and molecular data. This fungus was distinguished from other
Ganoderma species.
Ganoderma sichuanense was characterized by its distinctive substipitate to stipitate, flabellate to reniform, radially rugose pileus, laccate with a verrucose or tuberculose upper surface, pore surface yellowish when young, becoming brown or black when bruised, and small spores [
13]. Originally the basidiospores were described as (7.4–9.5 × 5–7) µm [
40], then updated to (7.8–10.4 × 5.2–6.4) µm [
13,
61], and (9–11.5 × 6.5–8) µm [
5]. The study basidiospores were 7.8–9.5 × 5.6–8.3 μm, which is in the range of the original report, which is not distinct from those of basidiospores found in other reports. Cao et al. [
4] stated that
G. sichuanense differs from
G. lingzhi as its sessile basidiocarps and smaller basidiospores (7.4–9.2 × 5–6.6) µm, with distinctive yellow context, thick dissepiments, absence of concentric growth zones in the context, basidiospore size, yellow pore surface, and presence of melanoid bands upon maturity [
4,
37].
Ganoderma sichuanense was yellowish-brown, with a dark brown eusporium bearing thick echinulae, overlaid by a hyaline myxosporium. However, among the Chinese
Ganoderma species,
G. flexipes,
G. multipileum,
G. sichuanense,
G. tropicum, and
G. tsugae are the most similar species to
G. lingzhi because they share a reddish-brown pileal surface, similar basidiospores, and cuticle cells [
4].
Ganoderma sinense J.D. Zhao, L.W. Hsu and X.Q. Zhang, Acta Mycologica Sinica. 19: 272 (1979) (
Figure 14)
= Ganoderma formosanum T.T. Chang and T. Chen. Transactions of the British Mycological Society. 82(4): 731 (1984)
Facesoffungi number: FoF 06253
Description: Basidiomes annual, stipitate, subdimidiate. Pileus 2–6 cm in length, 2–4 cm in width, and 0.3–1 cm thick. Pileus stipitate, subdimidiate to dimidiate, flabelliform, spathulate, umbonate, radial from the center extending to the margin, tough to break when dried, often thick at the center, slightly soft at the margin, light in weight when dried, and without woody. Pileus surface laccate, convex, some radial furrowed to furrowed, imbricate, incised, glossy, shiny, spathulate, shallow sulcate when fresh, umbonate or uneven, strongly laccate and glossy when mature, and weakly laccate where the new hyphae are in active development (margin), usually smooth layers at the center when young to age, irregularly ruptured crust overlying the context, and leathery when age when break. Pileus color usually homogenous with brownish-red (8C7–8C8) to reddish-brown (8D7–8D8) at the center toward stipe, extending brownish-red (9C8) from the center, slight to the margin when mature, usually reddish-brown (8E5–8E8) upper margin surface when old. Context up to 0.3–1 cm thick near stipe, dry, upper layer brownish-orange (6C8) when fresh, grayish-orange (5B5) at lower layers, with dark brown (8F7) when dried, soft and fibrous, covered with thin crust, some present woody, trimitic hyphal, hyaline, thin to thick-walled with simple septa, with branched. Tube 0.3–0.6 cm in length, brown (7D8). Stipe 4–16 cm in length, sub-cylindrical to cylindrical, almost stipitate with broadly and thick at the base, irregularly ruptured crust overlying, usually strongly laccate with brown (7D8) to dark brown (8F8) when mature, and dark brown (8F8) when old. Margin soft, some wavy, laccate when mature, weakly laccate when old, brownish-orange (6D8) when mature to old. Pore 4–6 in number per mm, subcircular to circular. Pore surface white (11A1) to yellowish-white (2A2) when mature, discolored when touched, brownish (6E7) to dark brown (6F7) when scratched or bruised.
Hyphal structure: Hyphal system trimitic, with clamp connections, usually light orange (5A5), orange (5A7), golden yellow (5B7–5B8), sometimes brownish-red (8C7) in KOH; generative hyphae 1.3–2.4 µm broad (n = 30), hyaline, thin-walled, with clamp connections; skeletal hyphae 3.1–5.2 µm broad (n = 30), usually hyaline, thick-walled, unbranched, and solid; binding hyphae 2.9–5.2 µm width (n = 30), thin to thick-walled, with branched, hymenial with sword-like apices in the context. Basidiospores mostly ellipsoid to broadly ellipsoid, with double walls, with size range of (9.5-)10.2–11.4–12.3(-13.1) × (7.0-)7.6–8.4–9.3(-10.2) μm ( = 11.2 × 8.5 μm, n = 50) μm, with Q = 1.28–1.36, L = 11.24 µm, W = 8.50 µm (including myxosporium), (8.3-)9.4–10.3–11.5(-12.3) × (6.1-)6.7–7.1–7.5(-8.0) μm ( = 10.2 × 7.2 μm, n = 50) μm, with Q = 1.41–1.47, L = 10.32 µm, W = 7.13 µm (excluding outer myxosporium), overlaid by hyaline, apically, short echinulae, truncate, some turgid vesicular appendix, inner wall echinulate, orange (5A7), deep orange (5A8, 5B8), orange (6B8), with brownish-orange (6B8), outer wall usually brownish-red (8C7–8C8) in 5% KOH.
Habitat: Solitary on decaying and living tree of Albizia mollis (Wall.) Boiv., living tree Quercus sp.
Specimens examined: CHINA, Yunnan Province, Baoshan, 25°09′35″ N, 99°09′49″ E, 1973 m elev., 11 November 2017, T. Luangharn, MLFU 19-2173.
Notes: Ganoderma sinense was described from China, characterized by a uniformly brown to dark brown context and slightly longitudinally crested basidiospores [
122]. This species was considered as a species with high phenotypic [
13,
36]. Our
G. sinense collection was obtained from Yunnan Province, China, and agrees well with the description of the holotype as described by Wang and Wu [
122]. Several reports have also illustrated
G. sinense from China [
13,
47,
123].
Ganoderma sinense is considered to have characteristics similar to
G. lucidum, while
G. sinense illustrates differences in macro- characteristics in its thin pileus, long stipes, and rarely branched skeletal hyphae with Bovista-type binding hyphae [
106], and these two different species are distinguished in reports [
130].
Ganoderma sinense is also reported to have similar characteristics with
G. formosanum; hence,
G. formosanum was treated as synonymous, and consequently, the earliest used valid name was
G. sinense [
122].
Ganoderma tsugae Murrill, Bulletin of the Torrey Botanical Club. 29; 601 (1902) (
Figure 15)
≡ Fomes tsugae (Murrill) Sacc. and D. Sacc., Sylloge Fungorum. 17: 123 (1905)
≡ Polyporus tsugae (Murrill) Overh.: 714 (1915)
= Polyporus metallicus Lloyd, Mycological Writings. 6 65): 1099 (1920)
Facesoffungi number: FoF 06254
Description: Basidiomes annual, subdimidiate, stipitate. Pileus 2–16 cm in length, 2–9 cm in width, and 0.5–3 cm thick at the base. Pileus stipitate, subdimidiate to dimidiate, flabelliform, spathulate, umbonate, concentrically sulcate zone, radial from the center extending to the margin, tough to break when dried, often thick at the center, slightly soft at the margin, light in weight when dried, with woody or corky when dried. Pileus surface laccate, convex, radial furrowed, imbricate, incised, glossy, shiny, spathulate, shallow sulcate, umbonate or uneven, strongly laccate and glossy when mature, and weakly laccate where the new hyphae are in active development (margin), usually smooth layers at the center when young to age, irregularly rugose, irregularly ruptured, thin crust overlying the context, tough to break when dried. Pileus color usually homogenous with brownish-red (8C7–8C8) to reddish-brown (8D7–8D8) at the center toward the stipe and margin surface when mature to old. Context up to 0.4–2.2 cm thick near the stipe, brownish-orange (7C7–7C8) to brown (7D8) on the upper layers, brownish-red (8C6) when dried, soft and fibrous, covered with thin crust, some present woody, dimitic hyphal, hyaline, thin-walled with simple septa, branched. Tube 0.3–1.6 cm in length, with dark brown (7F5). Stipe 4–10 cm in length, 2 cm thick, sub-cylindrical to cylindrical, almost stipitate and broad and thick at the base, irregularly ruptured crust overlying, usually strongly laccate with brown (7D8) to dark brown (8F8) when mature, usually dark brown (8F8) when old. Margin soft, some wavy, laccate when mature, and strong laccate when old, brownish-red (8C7–8C8) to reddish-brown (8D7–8D8) from mature to old. Pore 4–6 in number per mm, circular or angular. Pore surface yellowish-white (4A2) when present to yellowish-white (2A2) when mature, discolored when touched, brownish (6E7) when scratched or bruised.
Hyphal structure: Hyphal system trimitic, with clamp connections; generative hyphae 3.1–4.8 µm broad (n = 30), hyaline, thin-walled, with clamp connections; skeletal hyphae 3.1–6.8 µm broad (n = 30), usually hyaline, thick-walled, non-septate, unbranched, and solid; binding hyphae 3.9–5.0 µm width (n = 30), with walls varying in thickness, with many branches, some hymenial with sword-like apices in the context. Basidiospores mostly ellipsoid to broadly ellipsoid, with double walls, with size range of (9.7-)10.6–12.7–14.3(-15.8) × (7.3-)8.4–10.7–11.5(-12.4) μm ( = 12.7 × 10.5 μm, n = 50) μm, with Q = 1.18–1.24, L = 12.68 µm, W = 10.48 µm (including myxosporium), (8.3-)9.4–10.8–12.6(-13.1) × (6.1-)6.9–7.6–8.3(-9.2) μm ( = 10.7 × 7.6 μm, n = 50) μm, with Q = 1.36–1.45, L = 10.68 µm, W = 7.59 µm (excluding outer myxosporium), overlaid by hyaline, apically and short echinulae, truncate and turgid vesicular appendix, inner walled echinulate, brownish-orange (6C7–6C8), outer walled usually dark-brownish (6F7–6F8) in 5% KOH.
Habitat: Solitary, on decaying Quercus spp. tree.
Specimens examined: CHINA, Yunnan Province, Jinning District, 24°41′17″ N, 102°13′15″ E, 1973 m elev., 11 November 2017, JC. Xu, HKAS 97406.
Notes: Ganoderma tsugae has been treated as a synonym of
G. lucidum [
144,
145,
146]. This fungus is characterized by a laccate and concentric yellowish-red pileus, stipitate, fan-shaped, sulcated with a yellow margin, ovoid, verrucose, and truncated basidiospores.
G. tsugae is widely distributed across the USA [
35,
109]. The phylogenic analysis supported
G. tsugae as an independent species distinct from
G. lucidum, as it grows exclusively on conifers, especially on
Tsuga and
Abies species, while
G. lucidum inhabits mostly angiospermous trees [
76]. According to Loyd et al. [
35],
G. tsugae is similar to
G. oregonense as they share a distinctly white context tissue, rough basidiospores, and are predominately associated with conifers decay.
3.2.2. Taxonomy of Ganoderma from Laos
Ganoderma adspersum (Schulzer) Donk Proc. K. Ned. Akad. Wet., Ser. C, Biol. Med. Sci. 72(3): 273 (1969) (
Figure 16)
≡ Polyporus adspersus Schulzer, Flora.: 11 (1878)
= Polyporus linhartii Kalchbr., Fungi Hong. 252 (1884)
= Ganoderma europaeum Steyaert, Bulletin du Jardin Botanique de l'État à Bruxelles. 31: 70 (1961)
Facesoffungi number: FoF 06241
Description: Basidiomes annual, subdimidiate, sessile. Pileus 3–22 cm in length, 2–14 cm broad, and 1–4 cm thick at the base. Pileus sessile, perennial, subdimidiate to dimidiate, flabelliform, spathulate, umbonate, concentrically sulcate zone, somewhat round and plump when young, somewhat imbricate with flabelliform (fan-shaped) when seen from above, broadly attached, radial from the center extending to the margin, tough to break when dried, thick at the base, slightly soft at the margin when mature, light in weight with woody or corky when dried. Pileus surface non-laccate (dull), convex, radially furrowed, incised, spathulate, shallow sulcate, usually silky, soft, and smooth when young, and slippery surface when fresh, thick crust overlaying the context, a differentiated zone at the point of attachment, and tough to break when dried. Pileus color usually homogenous with reddish-orange (7A8) to brown (7D7–7D8) at the center when mature, golden yellow (5B7), brownish-orange (5C5–5C6, 6D5) when old toward the stipe and margin surface. Context up to 0.5–2.5 cm thick near stipe, brown (7D8) to brownish-red (8F8) when mature or dried, soft and fibrous, covered with hard and thick crust, woody when old, trimitic hyphal system present, hyaline, thin to thick-walled, branched. Tube 0.5–1.5 cm in length, usually homogenous with orange (5A7) to dark orange (5A8), reddish-orange (7A7–7A8), and grayish-red (8C7). Stipe 1–5 cm in length, 6 cm thick at the base, almost sessile or some short, stipitate, broad and thick at the base, usually non-laccate, and brown (7D8) to dark brown (8F8) when mature. Margin 0.5–4 cm thick, round, soft, brown (7D8) when mature to old, and usually concolourous with the pileus. Pore 4–6 in number per mm, subcircular to circular. Pore surface yellowish-white (2A2) when mature, discolored when touched, brown (7D8) when scratched or bruised.
Hyphal structure: Hyphal system di-trimitic, with clamp connections, orange (6A7) to deep orange (6A8), brownish-yellow (6C8) to brownish-orange (7C8); generative hyphae 1.4–2.8 µm broad (n = 30), hyaline, thin-walled, with clamp connections; skeletal hyphae 2.1–4.4 µm broad (n = 30), usually hyaline, thick-walled, and solid; binding hyphae 1.5–3.6 µm broad (n = 30), with walls varying in thickness, with many branches, some hymenial with sword-like apices in the context. Pileipellis a hymeniderm, brown (6E8) to dark brown (7F6), which is composed of apically clavate-like branched cells. Basidiospores mostly ellipsoid to broadly ellipsoid, sometimes ovoid, with double walls, with a size range of (6.9-)7.5–9.1–9.8(-10.6) × (4.7-)5.4–6.4–7.0(-7.7) μm ( = 9.1 × 6.4 μm, n = 50) μm, with Q = 1.38–1.45, L = 9.09 µm, W = 6.41 µm (including myxosporium), (5.6-)6.3–7.6–8.4(-9.2) × (4.2-)4.7–5.6–6.1(-6.6) μm ( = 7.6 × 5.7 μm, n = 50) μm, with Q = 1.35–1.40, L = 7.6 µm, W = 5.52 µm (excluding outer myxosporium), overlaid by hyaline, apically, and short, echinulae, a truncate and turgid vesicular appendix, light yellow (4A4–4A5), grayish-yellow (4B3–4B4) to brownish-orange (5C5–5C6), (5B8) of inner wall, outer wall usually yellowish-brown (5D8, 5E7–5E8) to brown (6D7–6D8) in 5% KOH.
Habitat: Solitary, near the roots of a living Mangifera indica tree.
Specimens examined: LAOS, Luang Namtha Province, 20°35′47″ N, 101°04′07″ E, 935 m elev., 20 June 2018, T. Luangharn, MFLU 19-2177.
Notes: Ganoderma adspersum was first reported by Donk [
147], who described it as
Polyporus adspersus Schulzer.
Ganoderma adspersum is characterized by a distinctive non-laccate, sessile, and applanate pileus.
Ganoderma adspersum is often confused with
G. applanatum,
G. australe, and
Polyporus [
148]. Ryvarden [
149] and Ryvarden and Gilbertson [
105] considered the correct name of
G. adspersum as a synonym of
G. australe, with
G. adspersum can be differentiated from
G. applanatum by its thicker at the base, and larger basidiospores, while
G. applanatum tends to emerge sharply at right angles [
105,
110], with molecular analysis also supporting the differentiation [
45,
76,
150,
151,
152,
153]. Our collections agree well with the description provided by Ryvarden and Gilbertson [
105].
Ganoderma australe (Fr.) Pat., Bull. Soc. mycol. Fr. 5(2, 3): 65 (1889) (
Figure 17)
Facesoffungi number: FoF 06242
Description: Basidiomes annual, perennial, sessile. Pileus 6–11 cm in length, 2–6.5 cm broad, and 0.8–2 cm thick. Pileus single, flabelliform, subdimidiate, spathulate, umbonate, sulcate, obtuse from the host, broadly attached, consistency hard and tough when mature, tough to break when dried, often thick at the center, slightly soft at the margin, and usually woody and corky when dried. Pileus surface corky, convex, furrowed, spathulate, mostly umbonate or uneven, usually non-laccate (dull) when mature to old, smooth layers when present, deep sulcate at the center, thick and hard crust, irregularly ruptured crust overlying the surface, presented dark brown (7F8) cracked crust when old, and tough to break when dried. Pileus color often homogeneous with pale red (7A5), reddish-orange (7A6–7A7), brown (7D8), to orange red (8B7–8B8) on the upper surface of the base closed to the margin when mature to old. Context up to 0.5–1.2 cm thick near stipe, fibrous, composed of coarse loose fibrils, brown (6D7–6D8), dark brown (6F7) to reddish-brown (8D8, 8D9), covered with thick crust, trimitic hyphal, thick-walled, dense with simple septa, typically with narrow lumen, flexuous, and many branches. Tube 0.4–1 cm in length, brown (7D8) to dark brown (6F8). Stipe sessile with broad attached. Margin white (4A1) when present to mature, soft and slippery when growing fresh, shallow sulcate at the margin, covered and blunt when old. Pore 4–6 in number per mm, subcircular to circular, sometimes angular. Pore surface initially white (4A1), slightly yellowish-white (3A2) when mature, brownish-red (8C4–8C5) when scratched, bruised, or discolored when touched.
Hyphal structure: Hyphal system trimitic, with clamp connections, usually brownish-orange (6C5–6C7) in KOH; generative hyphae 2.0–3.4 µm broad (n = 30), thin-walled, hyaline, tapering branches, with clamp connections; skeletal hyphae 3.1–4.5 µm broad (n = 30), usually thick-walled, sometimes branches, nearly solid; binding hyphae 2.5–3.9 µm width (n = 30), usually thick-walled, many branches, nearly solid, and hymenial with sword-like apices in the context. Pileipellis a hymeniderm, brown (7D8), composed of apically acanthus-like branched cells, dextrinoid. Basidiospores mostly ellipsoid to broadly ellipsoid, with double walls, with a size range of (6.5-)7.6–10.1–11.4(-12.5) × (5.9-)6.7–8.5–9.2(-10.3) μm ( = 7.2 × 5.9 μm, n = 50) μm, with Q = 1.19–1.26, L = 7.24 µm, W = 5.92 µm (including myxosporium), (5.1-)6.2–8.3–9.7(-10.9) × (4.4-)5.6–6.8–7.7(-8.8) μm ( = 8.2 × 6.8 μm, n = 50) μm, with Q = 1.17–1.26, L = 8.23 µm, W = 6.79 µm (excluding outer myxosporium), overlaid by hyaline, brown apically, bearing fine, distinct, short, echinulae, truncate, turgid vesicular appendix, inner wall light brown (6D4–6D5) to brown (7E7–7E8), and outer wall usually reddish-brown (8E5–8E6, 8F7) in 5% KOH.
Habitat: Solitary, on the decaying hardwood of Canarium spp. tree species.
Specimens examined: LAOS, Luang Namtha Province, 20°35′47″ N, 101°04′07″ E, 935 m elev., 20 June 2018, T. Luangharn, MFLU 19-2171.
Ganoderma gibbosum (Blume and T. Nees) Pat., Ann. Jard. Bot. Buitenzorg, suppl. 1: 114 (1897) (
Figure 18)
Facesoffungi number: FoF 06243
Description: Basidiomes annual or perennial, sessile, subflabellate, or subdimidiate. Pileus 2–16 cm in length, 2–9 cm broad, and 0.5–2.3 cm thick. Pileus conks, convex, imbricate, umbonate, uneven, ungulate, usually round when occurring, primordial, somewhat round and plump when young, flabelliform (fan-shaped) when seen from above, broadly attached when mature, thick at the base when mature. Pileus surface non-laccate, smooth when young, silky, soft, and slippery surface when fresh, furrowed on the surface with sulcate to undulating, somewhat spathulate to uneven, incised, compact, hard, and woody when older, covered with a tough crust (0.1–0.2 mm), usually dull and faded when mature to old, and some occurred the lined or cracked crust when older. Pileus color brownish-orange (5C5), reddish white (7A2) at the base, and homogenous with grayish-orange (6B3), brownish-orange (7C5), and light brown (6D4) toward the center of maturity fruiting bodies, white (6A1) at the margin, and usually the color changes to dark brown (8F8) upon touch, becoming grayish-red (8C4–8F6), reddish-brown (8E6) to dull red (10C3) when old. Context up to 0.3–1.3 cm thick, trimitic hyphal with clamp connections, hyaline, with walls varying in thickness, simple septate, composed of narrow, and sparse branches; generative hyphae 1.0–3.4 µm broad (n = 30), with walls varying in thickness, and hyaline; skeletal hyphae 4.0–6.4 µm broad (n = 30) with thick walls; binding hyphal 2.0–6.5 µm broad (n = 30). Hymenophore up to 3 mm in length, with reddish-brown (8D7). Tube layers 0.2–0.8 cm in length, up to 80–163 µm in width, and non-presented when young. Stipe almost sessile, broadly attached when present. Margin wavy, blunt, slippery when wet, thinner at the base and soft than the center, often white (8A1) from youth to maturity, and light brown (6D5) when old. Pore 4–7 in number per mm, when fresh, angular, subcircular to circular. Pore surface white (11A1) to orange white (6A2) when fresh, scratched or bruised, and discolored when touched.
Hyphal structure: Hyphal system trimitic, with clamp connections, usually reddish-brown (8D7–8D8); generative hyphae 1.0–3.8 μm broad (n = 30), with walls varying in thickness, hyaline, and unbranched; skeletal hyphae 4.2–6.4 μm broad (n = 30), light brown (7D6) to brown (7D8) in Melzer’s reagent with dextrinoid, usually thick-walled; binding hyphae 1.8–6.4 μm broad (n = 30), brown (6F8) to reddish-brown (8D8) in Melzer’s reagent, thick-walled, many branches, the generative and skeletal hyphae usually intertwined, mostly dark brown (6F7) near the tube layers; Bovista-type ligative hyphae, hymenial with sword-like apices in the context. Pileipellis a hymeniderm, brown (6D8) to light brown (6D6), composed of apically acanthus-like branched cells, dextrinoid. Basidiospores mostly ellipsoid to oblong ellipsoid or broadly ellipsoid, with double walls, with a size range of (4.4-)6.8–8.2–9.5(-10.2) × (3.6-)4.2–5.2–5.8(-6.5) μm ( = 8.3 × 5.4 μm, n = 50) μm, with Q = 1.49–1.56, L = 8.34 µm, W = 5.44 µm (including myxosporium), (3.6-)5.3–6.0–7.2(-8.3) × (2.8-)3.9–4.5–5.4(-6.2) μm ( = 6.2 × 4.5 μm, n = 50) μm, with Q = 1.34–1.40, L = 6.24 µm, W = 4.51 µm (excluding outer myxosporium), overlaid by hyaline, dextrinoid, echinulae, inner wall echinulate brown, light brown (6D6–6D8) to brown (6E8) in 5% KOH, and reddish-brown (8F6) to dark brown (8F8) in Melzer’s reagent. Basidia not seen.
Habitat: Occasionally on decaying wood of Pinus spp.
Specimens examined: LAOS, Luang Prabang, 19°51′51″ N, 102°11′39″ E, 589 m elev., 12 July 2018, T. Luangharn, MFLU 19-2190.
Ganoderma nasalaense Hapuar., Pheng. and K.D. Hyde, Mycosphere 10(1): 272 (2019)
Taxonomy and phylogeny analysis are shown in Hapuarachchi et al. [
30].
Notes: Ganoderma nasalaense is characterized by its duplex context, rigid basidiomes, purplish-black laccate crust, dark brown to gray on the upper pileus surface, brown tube layer, purplish-brown pore surface, trimitic hyphal system, and relatively large, broadly ellipsoid to ellipsoid light brown basidiospores (12.1–13.8 × 9.2–10.5) μm, truncate, with fine and short echinulae (10–12 × 6.5–7.5) µm, and a cuticle composed of strongly amyloid, clavate cells, usually with several irregular lobes or protuberances (30–80 × 3–10.5) µm. This fungus is solitary on decaying hardwood tree trunks in Huaphanh Province, Laos.
3.2.3. Taxonomy of Ganoderma from Myanmar
Ganoderma hoehnelianum Bres., Annales Mycologici. 10 (5): 502 (1912) (
Figure 19)
Facesoffungi number: FoF 06260
Description: Basidiomes annual or perennial, sessile. Pileus is 0.5–4 cm in length, 0.5–3 cm broad, and up to 0.5 cm thick at the base. Pileus is applanate, umbonate, sub-reniform to reniform, or subflabellate to glabrous from youth to maturity, small in size, obtuse from host, often with undefined concentric zones at the center that extend to the margin, thick at the center, slightly thin and soft at the margin, and tough to break when dried. Pileus surface shiny, smooth, and soft when young, frequently furrowed and shallowly sulcate on the upper surface, which is undulating and somewhat spathulate to uneven in maturity, covered by a thin crust, faded or weakly laccate when young, laccate when mature, and woody when old. Pileus color homogenous, reddish-brown (8E4–8E8) to dark brown orange (9F5–9F8) at the center when mature. Context up to 0.1–0.3 cm thick at the base, brown (7D7–7D8), reddish-brown (8E7–8E8), and dark brown (7F7), abundantly thick-walled, with clamp connections, subsolid hyphae, containing a fibrous pithy context. Tube layers hard and corky, branched, with clamped connections, often brown (7D7) to dark brown (7F8). Stipe almost sessile, broadly attached when present. Margin obtuse from the center, soft and smooth when young, laccate when mature, slightly wavy and slippery when wet, often yellowish-white (3A2) on the upper surface, and pale yellow (4A3) under the margin. Pores 4–6 in number per mm. Pore surface usually white (11A1) to light orange (5A4) when young, grayish-orange (6B4–6B6) when mature, turning light brown (7D5–7D6), brown (7D7), and reddish-brown (8D5–8D7) when scratched or bruised, becoming discolored when touched.
Hyphal structure: Hyphal system is dimitic, bearing clamp connections, hyaline, with walls varying in thickness with simple septa and some swollen differentiated zones at the point of attachment, composed of several narrow hyphae, and sparingly branched; generative hyphae (1.8-)2.4–3.2–3.7(-4.2) μm broad (n = 30) are thin-walled and hyaline; skeletal hyphae (3.3-)3.9–5.2–5.8(-6.2) μm broad (n = 30) have walls of varying thickness, sometimes subsolid; binding hyphae (2.3-)2.9–4.2–4.9(-5.6) μm broad (n = 30) are usually thick-walled with many branches, and appear alongside Bovista hyphae, which are usually present from orange (5A6–5A7) to deep orange (6A8) and thin-walled, as well as also reddish-brown (8D7–8D8, 9D8) to brownish-red (9C8) in Melzer’s reagent. Basidiospores are mostly ellipsoid and featuring several ovoid with double walls, (6.7-)7.5–9.8–11.8(-13.2) × (5.7-)6.4–7.8–9.6(-10.8) μm ( = 9.7 × 7.8 μm, n = 50) μm, with Q = 1.22–1.28, L = 9.73 µm, W = 7.78 µm (including myxosporium), (5.8-)6.4–8.5–9.3(-10.2) × (4.8-)5.3–6.1–6.8(-7.4) μm ( = 8.4 × 6.4 μm, n = 50) μm, with Q = 1.29–1.35, L = 8.43 µm, W = 6.41 µm (excluding outer myxosporium), overlaid by a hyaline, dextrinoid, and echinulate whose inner wall presents as brownish-orange (5C5–5C6) to brownish-yellow (5C7–5C8) and dark brown (6F8) to reddish-brown (8E7–8E8) in Melzer’s reagent.
Specimens examined: MYANMAR, Chin State, Tedim Township, 13 July 2019, P. E. Mortimer, MFLU 19-2168.
Notes: Ganoderma hoehnelianum was introduced by Bresadola in 1912 from Indonesia. Of distinctive note are its context color, basidiospore characteristics, and cuticular cells [
154]. Wang and Wu [
154] reported that the original Chinese
G. hoehnelianum was an earlier name for
G. shangsiense, and this fungus was also recorded as
G. shangsiense in China’s Hainan Province [
30,
91,
120,
121]. Our
G. hoehnelianum is first recorded from Myanmar, and its description is consistent with the descriptions provided by Wang and Wu [
154].
Ganoderma myanmarense Karunarathna, Mortimer, & Luangharn, sp. nov. (
Figure 20)
FacesofFungi number: FoF 06262
Index Fungorum number: IF 556794
Diagnosis: Ganoderma myanmarense is characterized by its shell-like pileus with strongly laccate appearance colored orange, golden yellow at center, extending reddish-yellow and yellow at margin, usually homogenous reddish-brown when mature, a white to light yellow that indicates active development on the margin of the pileal surface, a white pore surface when fresh, an orange, deep orange to reddish-orange context, and absence of melanoid bands.
Holotype: MYANMAR, Chin State, Tedim Township, on dead wood of Casternopsis sp., 13 July 2019, LT2019 (MFLU 19-2167).
Etymology: The species epithet “myanmarense” refers to the country Myanmar, where the holotype specimen was collected.
Description: Basidiomes annual or perennial, stipitate. Pileus up to 5–16 cm in length, 4–15 cm in width, and 1–2.5 cm thick at the base. Pileus shell-like (involute from the margin toward the center), sub-reniform to reniform or subflabellate to circular when viewed from above, often with undefined zones at the center that extend to the margin, with a thick center that is slightly soft at the margin, and tough to break when dried. Pileus surface shiny, silky, smooth, and soft when fresh, hard when old, furrowed, sulcate to undulating, somewhat spathulate to uneven, incised, faded or weakly laccate when young, strongly laccate on maturity or when old. Pileus color homogenous, orange (5A6–5A7, 6B7–6B8), golden yellow (5B6), and yellowish-red (8A7–8A8) at the center, slightly reddish-orange (7A7–7A8) and reddish-brown (8E8) where deeply sulcate, reddish-yellow (4A7) and yellow (2A6) at the margin. Context up to 0.6–1.5 cm thick at the base, with some areas thin-walled, thick-walled, along with subsolid hyphae, bearing clamp connections, and absence of melanoid bands. Tube layers woody and hard, usually thin-walled, frequently branched, with clamped connections, and often dark brown (7F7) when dried. Stipe up to 3–8 cm in length, up to 1.3–4 cm in width, centrally stipitate, nearly sub-cylindrical to cylindrical, concolorous with the pileus, often dark brown (9F7–9F8) to violet-brown (11F7–11F8), strongly laccate from maturity to old. Margin obtuse from the center, strongly laccate at the edge, occasionally wavy, slippery when wet, smoother, softer, and thinner, at the base, often yellow (2A6), and deep orange (5A8) to golden yellow (5B7–5B8) on maturity to old. Pores 4–6 in number in number per mm, (60–)80–125(–165) μm, angular, subcircular to circular. Pore surface usually white (11A1), turnins light brown (7D5), brown (7D7–7D8) to dark brown (7F6–7F8) when scratched or bruised, becomes discolored when touched.
Hyphal structure: Hyphal system trimitic, with walls of varying thickness, clamp connections present, simple septate hyphae, hyaline, narrow and sparingly branched; generative hyphae (1.2-)1.8–2.2–2.4(-2.7) μm broad (n = 30), hyaline, thin-walled; skeletal hyphae (1.8-)2.5–3.8–4.5(-5.0) μm broad (n = 30), abundant and thick-walled, sometimes subsolid; binding hyphae (1.5-)1.9–2.7–3.3(-4.3) μm broad (n = 30), usually thick-walled. Basidiospores (8.2-)9.3–11.6–12.5(-13.6) × (5.3-)6.0–7.1–7.8(-8.6) μm, (Qm = 1.7, Q = 1.2–2.4, including myxosporium) (n = 50), ( = 11.5 × 7.1 μm, n = 50) μm, with Q = 1.57–1.65, L = 11.52 µm, W = 7.13 µm (including myxosporium), (7.5-)8.3–9.1–10.4(-11.9) × (4.9-)5.4–6.2–6.8(-7.5) μm ( = 9.0 × 6.3 μm, n = 50) μm, with Q = 1.40–1.48, L = 9.04 µm, W = 6.27 µm (excluding outer myxosporium), mostly ellipsoid to broadly ellipsoid, or globose, double walls, overlaid by hyaline; exosporium (outer wall) smooth, hyaline, endosporium (inner wall) coarse and echinulate, with turgid vesicular appendix, and pale yellow inner wall that can also present in KOH, and yellowish-brown (5E6–5E7) to brown (6E6–6E7) in Melzer’s reagent, outer wall pale orange (6A3–6A4) to orange (6A6–6A7) in KOH, reddish-orange (7A8), grayish-red (7B6–7B8), and dark brown (7F7–7F8) in Melzer’s reagent.
Habitat: Solitary near the hardwood root of unknown tree species.
Additional specimens: MYANMAR, Chin State, Tedim Township, 13 July 2019, P. E. Mortimer, MFLU 19-2167 (holotype) and MFLU 19-2169 (paratype).
Ganoderma williamsianum Murrill, Bulletin of the Torrey Botanical Club 34: 478 (1907) (
Figure 21)
≡ Elfvingia williamsiana (Murrill) Imazeki, Bulletin of the Government Forest Experimental Station Meguro. 57: 106 (1952)
Facesoffungi number: FoF 06261
Description: Basidiomes annual or perennial, stipitate. Pileus 0.5–1.5 cm in length, 0.5–1 cm in width, and up to 0.5 cm thick at the base. Pileus sub-reniform to reniform, or subflabellate to circular when young, often with concentric zones at the center that extend to the margin, thick at the center, slightly soft at the margin, tough to break when dried. Pileus surface shiny, silky, and soft when young, generally furrowed, smooth, sulcate to undulating, somewhat spathulate to uneven when mature, hard and woody when old, incised on the surface, faded or weakly laccate when young, and usually laccate when mature. Pileus color usually homogenous, orange (5A7–5A8) to deep orange (5B7–5B8), slightly white (5A1), and yellowish-white (3A2, 4A2) at the margin when young. Context is up to 0.3–1.5 cm thick at the base, which is usually thick-walled, with abundant walls varying in thickness, subsolid hyphae containing a fibrous pithy context with clamp connections, and brown (7D8) melanoid bands when mature. The tube layers hard, frequently branched with clamp connections, and often dark brown (7F8). Stipe 1.5 cm in length, 1 cm in width when young, lateral to nearly dorsal, entrally stipitate, almost sub-cylindrical to cylindrical, concolorous with the pileus, thick when young, and often brownish-orange (7C6) to dark brown (9F7–9F8). Margin obtuse from the center, blunted when young, occasionally wavy, slippery when wet, smooth and soft when young (1.5 cm), and often white (3A1) to yellowish-white (3A2) when young. Pore surface white (11A1) to yellowish-white (3A2), turns light brown (7D5) to brown (7D7–7D8) when scratched or bruised, becomes discolored when touched.
Hyphal structure: Hyphal system trimitic, bearing clamp connections, hyaline, with walls of varying thickness, simple septate, swollen differentiated zones at the point of attachment, composed of several narrow hyphae, and sparingly branched; generative hyphae (1.6-)1.9–2.3–2.5(-2.6) μm broad (n = 30), thin-walled and hyaline; skeletal hyphae (3.1-)3.4–3.9–4.3(-4.8) μm broad (n = 30), with walls of varying thickness, with some subsolid; binding hyphae (1.6-)1.9–2.6–2.9(-3.4) μm broad (n = 30), usually thick-walled, appearing alongside Bovista hyphae with many branches, whose thick walls usually present as light orange (5A5), orange (5A6–5A7, 6B7–6B8) to deep orange (5A8, 6A8), and reddish-brown (8D7–8D8) to brownish-red (9C8) in Melzer’s reagent. Basidiospores not observed.
Specimens examined: MYANMAR, Chin State, Tedim Township, 13 July 2019, P. E. Mortimer, MFLU 19-2170.
Notes: Ganoderma williamsianum belongs to the group of laccate
Ganoderma. This fungus was originally reported in the Philippines and is easily recognized, with its small, dense, ungulate of pileus with pale, yellow pores, large spores, and a short skeletal [
2,
113]. Among the
Ganoderma species, there are some similarities between
G. williamsianum and
G. brownii, such as both having yellow pores [
109]; however,
G. brownii can be differentiated from
G. williamsianum by its dull pileus with a hard crust [
155], skeletal hyphae with occasional branching, and smaller basidiospores [
109,
110,
137,
155].
Ganodermawilliamsianum is the earliest valid name for
G. meijiangense [
154], containing similarities to
G. meijiangense, such as both having sessile, annual crust basidiomes, and white margin, but
G. williamsianum can be differentiated from
G. meijiangense by its distinguishing dark brown context, without any layer of black crust and a distinct cuticular composition [
61]. Our
G. williamsianum collection from Myanmar marks a new record, as it shares traits similar to Moncalvo and Ryvarden [
2], with sessile, annual crust-like basidiomes, and white margin, trimitic hyphal system bearing clamp connections, hyaline, and walls of varying thickness.
3.2.4. Taxonomy of Ganoderma from Thailand
Ganoderma adspersum (Schulzer) Donk Proc. K. Ned. Akad. Wet., Ser. C, Biol. Med. Sci. (
Figure 22)
Facesoffungi number: FoF 06241
Description: Basidiomes annual to perennial, applanate, subdimidiate. Pileus 2–14 cm in length, 2–7 cm in width, and 0.5–1.8 cm thick at base. Pileus subdimidiate to dimidiate, flabelliform, spathulate, umbonate, concentrically sulcate zone, sessile or short stipitate, distinctly contracted base, somewhat round and plump when young, somewhat imbricate when viewed from above, flabelliform (fan-shaped), usually broadly attached with radial from center extending to the margin, tough when break, thick at base, slightly soft at the margin when mature, light weight when dried, and woody and corky texture when dried. Pileus surface non-laccate (dull), convex, radial furrowed, incised, spathulate, shallow sulcate, usually silky, soft, smooth when young, and slippery surface when fresh, thick crust overlying the context, differentiated zone at the point of attachment, several layers thick, and leathery when broken. Pileus color usually homogenous with brown (7D7–7D8, 7E7–7E8, 8D8) at center toward stipe to margin surface when mature. Context up to 0.3–1.3 cm thick near the stipe, brown (7D8) to brownish-red (8F8) when mature and dried, soft and fibrous, covered with hard and thick crust, woody when old, trimitic hyphal, hyaline, with walls varying in thickness, with branches. Tube 0.2–1 cm in length, usually homogenous with orange (5A7) to dark orange (5A8), reddish-orange (7A7–7A8), and grayish-red (8C7). Stipe 1–3.8 cm in length, 3.5 cm thick at base, almost sessile, some shortly stipitate, broadly thick at base, usually non-laccate, brown (7D8) to dark brown (8F8) when mature. Margin round, soft, occurring brown (7D8) from mature to old, presented numerous undulations, and usually concolorous with the pileus. Pores 4–6 in number in number per mm, subcircular to circular. Pores surface yellowish-white (2A2) when mature, turns brown (7D8) when scratched or bruised.
Hyphal structure: Hyphal system di-trimitic, with clamp connections; generative hyphae 1.2–2.8 µm broad (n = 30), hyaline, thin-walled, with clamp connections; skeletal hyphae 2.0–4.4 µm broad (n = 30), usually hyaline, thick-walled, solid; binding hyphae 1.6–3.7 µm width (n = 30), with walls varying in thickness, many branches, without clamp connections, some hymenial with sword-like apices in the context. Basidiospores mostly ellipsoid, sometimes ovoid with double walls, (7.0-)7.7–9.0–9.9(-10.7) × (4.8-)5.3–6.5–7.1(-7.8) μm ( = 8.9 × 6.7 μm, n = 50) μm, with Q = 1.30–1.37, L = 8.91 µm, W = 6.69 µm (including myxosporium), (5.8-)6.4–7.7–8.6(-9.4) × (4.1-)4.5–5.6–6.0(-6.5) μm ( = 7.7 × 5.5 μm, n = 50) μm, with Q = 1.35–1.42, L = 7.69 µm, W = 5.52 µm (excluding outer myxosporium), overlaid by hyaline, apically and shortly echinulate, truncate and turgid vesicular appendix, inner wall brownish-yellow (5C7–5C8) and light brown (6D7–6D8), outer wall usually brownish-orange (7C8), brown (7D7–7D8) to dark brown (7E7) in 5% KOH.
Habitat: Solitary on decaying Pterocarpus sp.
Specimens examined: THAILAND, Kanchanaburi Province, 10 November 2018, T. Luangharn, MFLU 19-2178.
Ganoderma applanatum (Batsch) G.F. Atk., Annales Mycologici 6: 189 (1908) (
Figure 23)
Facesoffungi number: FoF 06249
Description: Basidiomes annual or perennial, subdimidiate to dimidiate, sessile. Pileus 2–12 cm in length, 2.5–6 cm in width, and 0.8–3.1 cm thick at the base. Pileus sessile, perennial, subdimidiate to dimidiate, subflabelliform to flabelliform, convex, imbricate, umbonate or uneven, rarely ungulate, with broadly attached when mature, thicker at base, slightly soft at margin when mature. Pileus surface furrowed, tuberculate to undulate, uneven, and incised when old, non-laccate (dull), compact and hard when mature, woody to corky texture when mature to old, covered with a thin crust (0.1–0.25 mm) overlies the pileus, and some cracked crust when old. Pileus color differentiated zone with peach red (7A4) to reddish-orange (7A6, 7B6) at base, toward to brownish-orange (7C6–7C8) with radius light brown (7D5) zone at center, and extend to dark brown (6F8) closed to active mycelium (margin) of maturity fruiting bodies. Context up to 0.5–2.3 cm thick, mostly brown (6E8) to dark brown (7F6–7F8) of cuticle cells, upper layers light orange (5A5), lower reddish-brown (8D8) with fibrous, some fibrous pithy context, usually separated layers of context tissue at base, and some occurred woody line. Tubes 0.8–2 cm in length, up to 70–160 µm in width, with sulcate at different levels. Stipe almost sessile (without stipe) with broadly attached when present, with differentiated zone at the point of attachment. Margin up to 1 cm thick, round, soft, often white (5A1), turning to light brown (6D4) and brown (6E8) when scratched or bruised, slippery when wet, soft when young, and thin than the center. Pore angular, subcircular, 4–6 in number per mm. Pore surface white (7A1) when fresh, quickly turning to light brown (7D6) to brown (7D8) when handled, scratched, and bruised.
Hyphal structure: Hyphal system trimitic; generative hyphae 0.5–2.8 µm ( = 2.4, n = 30) in diam, clamp connections, almost hyaline, abundant thin-walled and occasionally thick-walled, composed of narrow and sparingly branched; skeletal hyphae 2.7–4.9 µm broad (n = 30), usually thick-walled, hyaline, sometimes branched; binding hyphae 1.2–3.6 µm broad (n = 30), thick-walled, some branched, and intertwined the skeletal hyphae. Pileipellis a hymeniderm, brown (6E4), which composed of apically acanthus-like branched cells. Basidiospores mostly ellipsoid to broadly ellipsoid, sometimes subcircular with double walls, overlaid by hyaline, exosporium (outer wall) hyaline, endosporium (inner wall) coarse echinulate, with turgid vesicular appendix, truncate at the distal end, (10.0-)10.5–11.3–11.9(-12.4) × (7.2-)8.0–8.8–9.4(-10.2) μm, ( = 11.2 × 8.6 μm, n = 50) μm, with Q = 1.28–1.33, L = 11.23 µm, W = 8.62 µm (including myxosporium), (6.3-)7.4–8.5–9.3(-10.1) × (4.9-)6.3–7.4–8.7(-9.3) μm ( = 8.4 × 7.5 μm, n = 50) μm, with Q = 1.09–1.13, L = 8.42 µm, W = 7.58 µm (excluding outer myxosporium), brown (7D7–7D8) in KOH, and reddish-brown (8E6) to dark brown (8F4) in Melzer’s reagent. Basidia 14–20 × 8–10 μm, with four sterigmata.
Habitat: Solitary on rotten wood, dead trunks, and decaying stumps of Artocarpus spp., and Dipterocarpus spp.
Specimens examined: THAILAND, Kanchanaburi Province, 10 November 2018, T. Luangharn, MFLU 19-2175.
Ganoderma australe (Fr.) Pat., Bull. Soc. mycol. Fr. 5(2, 3): 65 (1889)
Facesoffungi number: FoF 02906
Characteristics follow Luangharn et al. [
59].
Description: Basidiomes applanate, spathulate. Pileus 2–7 cm in length, 1–15 cm in width, and 0.5–3 cm thick near the base. Pileus circular, applanate, spathulate, sometimes flabelliform clusters when young, dimidiate, semicircular at maturity, smooth when present. Pileus surface convex, furrow, glabrous, glossy, laccate, and consistently hard when fresh, and tough and light in weight when dry. Pileus color distinct concentric zones with light brown (6D4) to brown (6E8) at the center, slightly pale orange (5A3) to white (5A1) at the margin when young, becoming reddish-brown (8E6) to dark brown (9F8) at the center, dull red (8C4), and pale red (7A3) to white (8A1) at the margin at old age on the upper surface, brown to dark brown when dried, separated by a layer of context, usually brown to gray in winter or may fade as weathering destroys pigments on the pileus surface. Tube layers 0.2–1.2 cm in length, 50–180 µm in width, thick-walled. Stipe 3–5 cm in length, 1.5–3.5 cm in width, 1.3–3.3 cm thick, applanate, with umbo that slightly extended at the base. Margin up to 0.5–3 cm thick, thinner and lighter than the base, soft, round, pale yellow (4A3) to grayish brown (5D3) and reddish gray (8B2), changing to grayish-orange (6E3) when touched, thick toward the margin and downward toward the poreless marginal part of hymenophore. Pore angular, 4–6 in number in number per mm. Pore surface white (2A1) to pale yellow (2A3) in growing specimen, immediately discolored when bruised, cream to grayish brown (5D3) when fresh, with brownish gray (5C2) to brown (4D7) when dried.
Hyphal structure: Hyphal system di-trimitic, contextual generative hyphae, binding, and skeletal hyphae, generative hyphae, 1–5 µm broad, with clamp connections, hyaline, thin-walled; binding hyphae 2–6 µm broad, thin to thick-walled, branched, with clamp connections; skeletal hyphae, hyaline, pale brown to brown, thick-walled, 2–7 µm broad. Basidiospores (6.4)6.9–9.3–10.4(11.1) × (5.8)6.4–7.9–8.8(9.7) μm, ( = 9.4 × 7.7 μm, n = 50), with Q = 1.20–1.21, L = 9.63 µm, W = 7.96 µm (including myxosporium), (5.4)6.3–7.0–8.1(8.9) × (3.4)3.8–5.9–6.5(7.3) μm, ( = 7.1 × 5.8 μm, n = 50) μm, with Q = 1.19–1.25, L = 7.29 µm, W = 5.93 µm (excluding outer myxosporium), reddish-brown, mostly broadly ellipsoid at maturity, some distinct tapering at the distal end, truncate, double wall, thick-walled inner endosporium. Basidia not observed. Cultures characteristics turned white after incubation at 30 °C for 10 days. Odor distinctive when dried.
Habitat: Solitary on hardwood Shorea robtusa, or rotten wood, dead trunks, decaying hardwood, decaying stumps, and occasionally occurring on standing trees or trunks of many broad-leaf trees.
Specimens examined: THAILAND, Chiang Mai Province, August 2012; T. Luangharn, MFLU 13-0534, MFLUCC 12-0527.
Ganoderma casuarinicola J.H. Xing, B.K. Cui and Y.C. Dai., MycoKeys 34: 93–108 (2018)
Facesoffungi number: FoF 06130
Taxonomy and phylogeny analysis are shown in Luangharn et al. [
31]
Notes:
Ganoderma casuarinicola was collected on a
Pinus kesiya stump in a pine forest. This fungus is distinctive by its strongly laccate, shallow sulcate, reddish-brown pileus surface, lateral stipe, white pore surface, and brown context. Thai
G. casuarinicola shows its annual, applanate to dimidiate, 3–16 cm long and 1.5–3 cm wide pileus, larger than Guangdong collection. Our
G. casuarinicola collections show longer tubes of 6–14 mm, while the tubes of the Guangdong collection are 9 mm long; however, our collections reveal a thinner margin (0.8–1.2 cm thick) than the Guangdong collection (2 cm thick). However, the type of
G. casuarinicola from the Guangdong collection does not have the melanoid band [
37], while our collection features a dark brown melanoid band. Micro- characteristics are dense light brown to brown context layers; walls of varying thickness in generative hyphae; thin-walled binding hyphae; and a thick-walled skeletal. Our
G. casuarinicola collection has mostly distinctive yellowish-brown basidiospores, with a smaller size range of (8.7)10.8–13.5(14.4) × (6.6)7.6–8.9(9.8) μm than the type of
G. casuarinicola (8.3-)9.0–10.2(-11.5) × (4.5-)5.0–6.0(-7.0) µm (including myxosporium).
Ganoderma ellipsoideum Hapuar., T.C. Wen and K.D. Hyde, Mycosphere. 9(5): 951 (2018) (
Figure 24)
Facesoffungi number: FoF 06255
Description: Basidiomes annual, sessile. Pileus 3–9 cm in length, 2.2–5 cm in width, and 1.5–3.5 cm thick at the base. Pileus annual, convex, imbricate, sessile, umbonate, uneven, ungulate, subflabellate or subdimidiate, somewhat imbricate, when seen from above flabelliform (fan-shaped), usually round, when present primordial, somewhat round and plump when present, broadly attached, thick at base, slightly soft at margin when mature. Pileus surface non-laccate (dull), furrowed, incised, sulcate, smooth when young, undulating on the upper surface, somewhat spathulate to uneven, covered by a thin and hard crust (0.1–0.4 mm), and woody when older. Pileus color usually homogenous with reddish-brown (9E7–9E8) to dark brown (9F7–9F8) at the base at the center, extending white (4A1) to brown (7E7) on the upper margin surface of mature fruiting bodies. Context up to 0.5–2.5 cm thick, compact and hard, trimitic hyphal, with clamp connections, hyaline, thin-walled with simple septa, sparingly branched; generative hyphae 1.2–3.7 µm broad (n = 30), hyaline, simple septate, with clamp connections; skeletal hyphae 1.8–4.2 µm broad (n = 30), usually thick-walled, unbranched; binding hyphal 2.0–4.8 µm width (n = 30) with sparingly branched, thick-walled, without clamp connection. Hymenophore usually brown (7D8) to reddish-brown (8E8). Tube layers 0.5–2.2 cm in length. Margin blunt-edged, wavy, slippery when young, and often white (8A1), where the new hyphae are in active development when young to mature. Pore 4–6 in number per mm, angular, subcircular to circular. Pore surface white (11A1) to pale yellow (2A3), turns brown (7E7) to dark brown (7F7–7F8) when scratched or bruised.
Hyphal structure: Hyphal system trimitic, with clamp connections, with brown (7E7); generative hyphae (1.2-)1.6–2.4–3.0(-3.8) μm broad (n = 30), branched, thin-walled, hyaline, with grayish-yellow (4B5) in KOH; skeletal hyphae (1.7-)3.1–3.8–4.3(-4.8) μm broad (n = 30), dextrinoid, abundant thick-walled, with unbranched; binding hyphae (2.1-)3.1–3.7–4.2(-4.6) μm broad (n = 30), thick-walled, frequently branched, usually intertwined the generative and skeletal hyphae, mostly brown (7E7) to dark brown (7F5) near the tube layers; Bovista-type ligative hyphae, hymenial with sword-like apices in the context. Basidiospores mostly ellipsoid with double walls, (4.8-)5.3–6.6–7.2(-7.7) × (3.1-)3.5–4.3–5.0(-5.4) μm ( = 6.8 × 4.5 μm, n = 50) μm, with Q = 1.34–1.43, L = 6.28 µm, W = 4.52 µm (including myxosporium), (3.6-)4.1–5.5–6.0(-6.4) × (1.7-)2.1–2.8–3.3(-3.7) μm ( = 5.49 × 2.83 μm, n = 50) μm, with Q = 1.83–1.92, L = 5.50 µm, W = 2.94 µm (including myxosporium), overlaid by hyaline, dextrinoid, echinulae, inner wall echinulate brownish-yellow (5C7–5C8) to brown (7D7–7D8) in 5% KOH. Basidia not seen.
Habitat: Solitary on rotten wood of Acacia sp.
Specimens examined: THAILAND, Chiang Mai Province, Mae Taeng, Mushroom Research Centre, 19°07′200″ N, 98°41′44″ E, 652 elev., 14 June 2019, P. E. Mortimer, MFLU 19-2221.
Ganoderma gibbosum (Blume and T. Nees) Pat., Ann. Jard. Bot. Buitenzorg, suppl. 1: 114 (1897) (
Figure 25)
Facesoffungi number: FoF 06246
Description: Basidiomes annual or perennial, sessile. Pileus 2–12 cm in length, 2–8 cm in width, and 0.5–2.8 cm thick. Pileus sessile (without stipe), subflabellate or subdimidiate, convex, imbricate, umbonate, uneven, round when occurring, primordial, round and plump when youth, with broadly attached when mature, and thicker at base slightly soft at margin when mature. Pileus surface non-laccate (dull), smooth, soft, slightly dull and faded when mature to old, usually silky when young, furrowed, tuberculate to undulate, uneven, and incised when mature, compact and hard when mature, woody when mature to old, covered with a thick crust, and some cracked crust when old. Pileus color light orange (6A4–6A5) to grayish orange (6B3–6B5) at base toward the center of mature fruiting bodies become grayish-green (30E2–30E7) on the upper surface when mature to old. Context up to 0.4–2.3 cm thick, tri-dimitic hyphal, with clamps connections, brownish-orange (7C7–7C8) to reddish-brown (8D7–8D8), mostly dark brown (7F7) near the tube layers, Bovista-type ligative hyphae, hymenial with sword-like apices in the context. Tube 0.1–0.6 cm long, up to 80–160 µm in width, and sulcate at different levels. Stipe almost sessile with broadly attached when present. Margin wavy, blunt-edged, slippery when wet, soft when young, thinner than the base and softer than the center, and often white (8A1) when young to mature. Pore angular, subcircular, 4–6 in number per mm in fresh. Pore surface initial white (7A1), turns light brown (7D6) to brown (7D8) when scratched or bruised, with a slippery surface when fresh.
Hyphal structure: Hyphal system trimitric hyaline, with walls varying in thickness with simple septa, composed of narrow and sparingly branched; generative hyphae 1.3–4.6 µm broad (n = 30), thin-walled and hyaline hyphae; skeletal hyphae 4.0–7.3 µm broad (n = 30), thick-walled and hyaline hyphae; binding hyphae 2.8–6.3 µm broad (n = 30), usually with walls varying in thickness. Pileipellis a hymeniderm, brownish-orange (6C8) to brown (6D8), composed of apically acanthus-like branched cells, dextrinoid. Basidiospores (4.2-)6.5–8.1–10.3(-11.2) × (3.9-)4.8–4.6–5.7(-6.4) μm ( = 8.2 × 4.7 μm, n = 50) μm, with Q = 1.70–1.76, L = 8.38 µm, W = 4.85 µm (including myxosporium), (4.5–)6.1–7.2–8.1(-9.2) × (3.9-)4.5–5.3–5.2(-6.9) μm ( = 7.4 × 5.4 μm, n = 50), with Q = 1.31–1.38, L = 7.53 µm, W = 5.60 µm (excluding outer myxosporium), ellipsoid or some globose with double walls, overlaid by hyaline, dextrinoid, distinct echinulate, inner wall echinulate brown, light brown (6D4) to brown (6E8) in 5% KOH. Basidia not seen.
Habitat: Solitary on standing trees of Dendrocalamus strictus.
Specimens examined: THAILAND, Chiang Rai Province, 19°48′20″ N, 100°04′19″ E, 680 m elev., October 2017, T. Luangharn, MFLU 19-2176.
Ganoderma lucidum (Curtis) P. Karst., Revue Mycologique Toulouse. 3(9): 17 (1881) (
Figure 26)
Facesoffungi number: FoF 06250
Description: Basidiomes annual or perennial, sub-reniform to reniform, stipitate. Pileus up to 2–6 cm in length, 1.5–3.0 cm in width, and 0.8–2.0 cm thick at the base. Pileus stipitate, sub-reniform to reniform, undefined imbricate, irregular, some laterally, and flabelliform with a contracted, concentrically sulcate zone, irregularly ruptured crust overlying the context, radial from center extending to the margin, tough when broken, often thick at center slightly soft at margin, and leathery when age when broken. Pileus surface smooth layer at center from young to old, usually furrowed, incised, undulate to sulcate, somewhat spathulate to uneven, some woody or corky texture when old, weakly laccate when present, strongly laccate and glossy when mature, and usually weakly laccate where the new hyphae are in active development (margin). Pileus color usually brownish-red (8C7–8C8) at the center, slight to reddish-orange (7B7–7B8), and orange (6A7–6A8) on the upper pileus surface. Context up to 0.4–1.4 cm thick at base, abundant thick-walled, subsolid hyphae, concentric lines of various shades, bearing clamp connections, light brown (6D6) to brown (6D8, 6E8), with dark brown (7F7) melanoid bands. Tube usually hard, brown (7D7) to dark brown (7F7). Stipe up to 8–16 cm in length, up to 0.8–1.5 cm in width, eccentric stipe, cylindrical to slightly flattened, laccate, and reddish-brown (8D7–8D8, 8E7–8E8) from mature to old. Margin often 0.5–1.3 cm, orange (6A7–6A8) upper surface, and reddish-yellow (4A8) under surface, thinner than the base, and softer than the center. Pore 4–6 in number per mm, (70–)110–145(–160) μm, subcircular to circular, sometimes angular. Pore surface white (11A1) when present, yellowish-white (2A3) to light brown (7D6) from young to mature, turning brown (7D7–7D8) to dark brown (6F6) when scratched or bruised.
Hyphal structure: Hyphal system trimitic, with clamp connections, hyaline, walls varying in thickness with simple septa, sparingly branched, swollen by melanoid bands, usually brownish-orange (6C7–6C8) to brown (6D7–6D8) in KOH; generative hyphae up to (1.8–)2.0–2.3–2.7(–3.0) μm broad (n = 30), usually thin-walled, some thick-walled, with clamp connections, branched, and almost hyaline; skeletal hyphae (3.2–)4.3–5.4–6.1(–6.8) μm broad (n = 30), usually thick-walled with clamp connections, with unbranched; binding hyphae (2.4-)2.9–4.4–5.0(-5.9) μm broad (n = 30), walls usually varying in thickness with abundant branched and present the melanoid bands. Basidiospores ellipsoid, some subglobose to globose with double walls, with a truncate apex, size range of (8.2-)8.8–9.8–10.5 (-11.4) × (5.8-)6.2–6.8–7.5(-8.2) μm, ( = 10.0 × 6.9 μm, n = 50) μm, with Q = 1.51–1.57, L = 10.65 µm, W = 6.92 µm, (including myxosporium), (6.3-)7.1–7.6–8.2(-8.4) × (4.8-)5.4–5.7–6.1(-6.5) μm, ( = 7.5 × 5.8 μm, n = 50) μm, with Q = 1.26–1.31, L = 7.57 µm, W = 5.89 µm (excluding outer myxosporium) (n = 50), brownish-orange (6C8, 6D8) to brown (6E5) of endosporium (inner wall) with brown (7E7–7E8) exosporium (outer wall) in Congo red, brownish-orange (6C8) in 5% KOH, and yellowish-brown (5D8) in Melzer’s reagent, mostly overlaid by hyaline, coarse echinulate, hyaline turgid vesicular appendix.
Habitat: Solitary on decaying hardwood of Dendrocalamus strictus in dry evergreen forest.
Specimens examined: THAILAND, Chiang Rai Province, 19°48′20″ N, 100°04′19″ E, 680 m elev., 15 October 2017, T. Luangharn, MFLU 19-2162.
Ganoderma multipileum Ding Hou. (1950) (
Figure 27)
Facesoffungi number: FoF 06256
Description: Basidiomes annual, stipitate. Pileus up to 6–11 cm in length, 4–9 cm in width, and 1–3 cm thick at base. Pileus sub-reniform to reniform, subflabellate to flabellate, concentrically sulcate zone, fleshed at center slightly to margin, radial or branched from center extend to the margin when seen from above, tough when break, often thick at center slightly soft at margin, and leathery when age when break. Pileus surface weakly laccate to strong laccate at center when mature to age, and faded or week laccate at active mycelial (margin), smooth, irregularly ruptured crust overlying the context, some woody or corky texture when old, usually furrowed, incised, sulcate to undulating, and somewhat spathulate to uneven on the surface. Pileus color usually homogenous brownish-red (8C7–8C8) at the base, slight yellowish-red (8A7–8A8, 8B7–8B8) at center, and light brown (7D5) to brown (7D7–7D8) on the upper margin surface. Context up to 0.3–1.2 cm thick at the base, with walls varying in thickness, subsolid hyphae, bearing clamp connections, usually light brown (6D4–6D6) to brown (6D8, 6E8) of hyphae, and dark brown (7F7) melanoid bands in KOH. Tube hard, usually thin-thick-walled, often with brown (7D7) to dark brown (7F7). Stipe up to 4–9 cm in length, up to 1–2.5 cm in width, almost eccentric, sub-cylindrical to cylindrical, plump, strong laccate, often homogeneous with red (9A6) to brownish-red (9C6), and dark red (10C7) when mature to old. Margin often white (4A1) to pale yellow (4A4) where the new hyphae are in active development, light brown (7D5) to brown (7D7) when bruised, strong laccate, wavy, slippery when wet, softer, thin than the base, and soft than the center. Pore angular, 4–6 in number per mm, (99-)120–154(-170) μm in diam, subcircular to circular. Pore surface white (11A1) when present, pale yellow (4A3) to orange white (5A2) when young to mature, light brown (6D4) with age, turning to light brown (7D6), brown (7D7–7D8) when dried or scratched and bruised.
Hyphal structure: Hyphal system trimitic, with clamp connections, hyaline, walls varying in thickness with simple septa, sparingly branched, usually light orange (5A4–5A5) to orange (5A6–5A7) in KOH; generative hyphae up to (1.6-)2.2–3.5–4.6(-5.2) μm broad (n = 30), usually thick-walled, unbranched, flexuous, and almost hyaline; skeletal hyphae (3.8-)4.2–5.1–5.9(-6.7) μm broad (n = 30), usually thick-walled, unbranched, sometimes subsolid; binding hyphae (1.4-)2.2–3.6–4.4(-5) μm broad (n = 30), usually walls varying in thickness, with flexuous, abundant branched, with Bovista-type binding hyphae, and occurred the melanoid bands. Basidiospores mostly ellipsoid, some ovoide, truncate at maturity, with double walls, (7.6-)8.8–11.7–12.4(-13.1) × (4.9-)5.3–6.1–6.9(-7.4) μm, ( = 11.8 × 6.3 μm, n = 50) μm, with Q = 1.84–1.89, L = 11.84 µm, W = 6.32 µm (including myxosporium), (5.9-)6.9–8.2–9.3(-10.6) × (4.3-)4.9–5.6–6.0(-6.5) μm, ( = 8.3 × 5.8 μm, n = 50) μm, with Q = 1.40–1.46, L = 8.36 µm, W = 5.85 µm (excluding outer myxosporium), mostly overlaid by hyaline, brownish-orange (6C8), (6D8) of exosporium (outer wall), endosporium (inner wall) coarse echinulae, with hyaline turgid vesicular appendix, with orange (6A7), (6B7) in KOH.
Habitat: Solitary on decaying stump of Pinus merkusii.
Specimens examined: THAILAND, Prachuap Khiri Khan Province, 12°08′52″ N, 99°45′41″ E, 491 m elev., 26 June 2018, T. Luangharn, MFLU 19-2166.
Notes: Ganoderma multipileum was originally reported from Taiwan, PRC [
44], which was presented over half a century ago from Taiwan, PRC [
39]. This fungus was the earliest valid name with
G. lucidum from tropical Asia [
44]; however, Wang et al. [
33] verified that
G. multipileum is the correct name for this tropical fungus. This fungus is a distinctive form with its laccate to strong laccate pileus, stipitate, rarely sessile, irregularly ruptured crust overlying the context, yellow-brown to dark brown context, cream pore surface, flattened or sub-cylindrical, lateral, horizontally lateral, with ovoid to ellipsoid basidiospores (7.6–13.5 × 5.5–7.5 μm) (with myxosporium), 6.5–10.5 × 4.5–6.5 μm (without myxosporium)), mostly truncate, brown, with a dark brown [
33]. Some researchers have shown in their phylogenies that
G. tropicum is similar to
G. multipileum [
4,
5], which is considered most resemble
G. tropicum in morphology and habitat even though they are distinct species [
33] and
G. flexipes [
4,
76,
129]. Our
G. multiplicatum was collected from Thailand, are similar to the original description by Wang et al. [
33] by its showed laccate to strong laccate pileus, radial or branched from center extends to the margin, with ellipsoid basidiospores.
Ganoderma orbiforme (Fr.) Ryvarden, Mycologia. 92(1): 187 (2000)
Figure 28)
≡ Polyporus orbiformis Fr., Epicrisis Systematis Mycologici.: 463 (1838)
≡ Fomes orbiformis (Fr.) Cooke, Grevillea. 14 (69): 18 (1885)
≡ Scindalma orbiforme (Fr.) Kuntze, Revisio generum plantarum. 3(2): 519 (1898)
≡ Ganoderma lucidum var. orbiformis (Fr.) Rick, Iheringia. 7: 201 (1960)
≡ Ganoderma orbiformum (Fr.) Ryvarden (2000)
= Ganoderma mastoporum (Lév.) Pat., Bulletin de la Société Mycologique de France. 5: 71 (1889)
= Ganoderma fornicatum (Fr.) Pat., Bulletin de la Société Mycologique de France. 5: 71 (1889)
= Ganoderma boninense Pat., Bulletin de la Société Mycologique de France 5: 72 (1889)
= Ganoderma subtornatum Murrill, Bulletin of the Torrey Botanical Club. 34: 477 (1907)
= Ganoderma cupreum (Cooke) Bres., Annales Mycologici. 9: 268 (1911)
= Ganoderma densizonatum J.D. Zhao and X.Q. Zhang, Acta mycol. sin.: 86 (1986)
= Ganoderma limushanense J.D. Zhao and X.Q. Zhang, Acta mycol. sin.: 219 (1986)
Facesoffungi number: FoF 06257
Description: Basidiomes annual or perennial, sessile. Pileus 4–11 cm in length, 3–6 cm in width, and 1–2.4 cm thick at base. Pileus sessile, flabelliform or spathulate, convex, imbricate, umbonate, uneven, ungulate, sub-reniform, sub-orbicular, subdimidiate, obtuse from host, broadly attached, somewhat imbricate, thicker at base, slightly soft at margin when mature. Pileus color usually homogenous with grayish-orange (6B3–6B6), brownish-orange (6C5–6C6), and brown (6D7–6D8) at base extending to the margin of maturity to old. Pileus surface partly non-laccate (dull) to weakly laccate, faded texture when old, furrowed, incised, sulcate, undulating, and somewhat spathulate to uneven on the upper surface, silky and soft when fresh, woody when mature to older, and covered with compact and hard crust (0.1–0.25 mm). Context up to 0.4–1.0 cm thick, trimitic hyphae with clamp connections, hyaline, walls varying in thickness with simple septa, sparingly branched. Hymenophore brown (6D7–6D8) to reddish-brown (8D7). Tube layers 0.3–1.2 cm in length, light brown (6D4–6D5) to brown (7D7). Stipe almost sessile with broadly attached when present, with brownish-orange (6C8). Margin soft, wavy, blunt-edged, slippery when young, with grayish-orange (6B5–6B6) to brownish-orange (6C8) when old. Pore 4–6 in number per mm, circular or subcircular, or angular. Pore surface white (11A1), when present, turns light brown (7D5) to brown (7D7–7D8) when scratched or bruised, becoming discolored when touched.
Hyphal structure: Hyphal system trimitic, with clamp connections, usually orange (6A6–6A8) to brownish-orange (6C7–6C8) in KOH; generative hyphae 1.4–5.0 µm broad (n = 30), hyaline, thin-walled with clamp connections; skeletal hyphae 2.6–5.6 µm broad (n = 30), usually thick-walled, unbranched; binding hyphae 1.2–4.8 µm width (n = 30), usually walls varying in thickness, many branches, hymenial with sword-like apices in the context. Basidiospores mostly ellipsoid to oblong ellipsoid or broadly ellipsoid, with double walls, (7.1-)7.9–9.4–11.2(-11.8) × (5.2-)5.97–6.7–7.1(-7.7) μm ( = 9.6 × 6.8 μm, n = 50) μm, with Q = 1.38–1.44, L = 9.63 µm, W = 6.82 µm (including myxosporium), (6.4-)5.2–6.0–6.7(-10.7) × (3.9-)4.5–5.3–6.1(-6.6) μm ( = 6.2 × 5.1 μm, n = 50) μm, with Q = 1.20–1.27, L = 6.28 µm, W = 5.10 µm (excluding outer myxosporium), overlaid by hyaline, echinulate, inner wall echinulate brown, some turgid vesicular appendix, light yellow (4A5) to reddish-yellow (4B7–4B7) in 5% KOH. Basidia not seen.
Habitat: solitary, on the living tree of Albizia mollis in deciduous forest, and living tree of Indochinese spp.
Specimens examined: THAILAND, Chiang Rai Province, 19°48′20″ N, 100°04′19″ E, 680 m elev., 21 October 2017, T. Luangharn, MFLU 17-1933.
Notes: Ganoderma orbiforme (Fr.) Ryvarden was first described as Polyporus orbiformis, with the original specimen from the tropical region of Guinea in Africa [
136]. The fungus was characterized by its distinctive weakly laccate surface or some dull surface areas, brown context, brown pore surface, and brown tube layer, and ellipsoid or ovoid basidiospore. G
anoderma cupreum, G. densizonatum, G. fornicatum, G. limushanense,
G.
multiplicatum, and G. subtornatum are similar to
G.
orbiforme; however, taxonomy and molecular analysis treated those taxa as the earliest valid names for
G.
orbiforme [
7,
156]. This fungus has been recorded from China, Laos, Myanmar, and Thailand [
30], with our collection is also the collection of
G.
orbiforme from Thailand. Our collection from Thailand agrees well with the description by Ryvarden [
136] and Wang et al. [
156] reported that this fungus posed rigid basidiomes, purplish-black laccate crust, brown pore surface, and tube layer, with ellipsoid or ovoid basidiospores, fine and short echinulate, clavate cells usually with several irregular lobes.
Ganoderma philippii Bres. and Henn. ex Sacc. Bres, Iconographia mycological. 21: 1014, t. 1014 (1932) (
Figure 29)
≡ Fomes philippii Bres. and Henn. ex Sacc., Sylloge Fungorum. 9: 180 (1891)
≡ Scindalma philippii (Bres. and Henn. ex Sacc.) Kuntze, Revisio generum plantarum. 3(2): 519 (1898)
= Fomes pseudoferreus Wakef., Bulletin of Miscellaneous Informations of the Royal Botanical Gardens Kew. 1918: 208 (1918)
Facesoffungi number: FoF 06258
Description: Basidiomes annual, sessile. Pileus 2–9 cm in length, 2–5 cm in width, and 0.4–3.8 cm thick at the base. Pileus annual, convex, sessile, umbonate, ungulate, usually round when present, primordial, plump when present, broadly attached and thick at base, slightly soft at margin. Pileus surface non-laccate to weakly laccate, furrowed, incised, sulcate, smooth when young, undulating on the upper surface, spathulate to uneven, covered with a crust (0.1–0.3 mm), cracked crust when old, and woody when old. Pileus color usually homogenous, brown (7E7–7E8) at base at the center, extending to brownish-orange (6C8), and white (4A1) when present, slightly yellowish-white (2A2) on the upper margin surface of mature fruiting bodies. Context consists of trimitic hyphae, up to 0.3–1.8 cm thick, sparingly branched, walls varying in thickness, compact and hard, with clamp connections, hyaline; generative hyphae 1.8–3.3 µm in width (n = 30), brownish-yellow (5C7–5C8), thin-walled, simple septa, hyaline, with clamp connections; skeletal hyphae 2.8–4.9 µm broad (n = 30), brownish-yellow (5C7–5C8), usually thick-walled, unbranched; binding hyphae 2.0–4.2 µm width (n = 30), brownish-yellow (5C7–5C8) to brownish-orange (6C7–6C8), sparingly branched, walls varying in thickness, and without clamp connections. Hymenophore heterogeneous, brown (7C7–7C8, 7D8), and melanoid band present when mature. Tube layers 0.3–3.8 cm in length. Margin blunt-edged, wavy, and often white (8A1) where the new hyphae are in active development to yellowish-white (2A2) from young to mature. Pore 4–7 in number per mm, subcircular to circular. Pore surface white (11A1) when present, turning brown (7E7) to dark brown (7F7–7F8) when scratched or bruised.
Hyphal structure: Hyphal system trimitic, with clamp connections, usually brown (7E7); generative hyphae (1.7-)2.1–2.5–2.9(-3.3) μm broad (n = 30), branched, thin-walled, hyaline, unbranched, grayish-yellow (4B5) in KOH, with clamp connections; skeletal hyphae (2.9-)3.4–4.0–4.4(-5.0) μm broad (n = 30), dextrinoid, abundant thick-walled, with unbranched; binding hyphae (2.1-)2.8–3.3–3.8(-4.3) μm broad (n = 30), walls varying in thickness, frequently branched, generative and skeletal hyphae usually intertwined, mostly brown (7E7) to dark brown (7F5) near the tube layer; Bovista-type ligative hyphae, hymenial with sword-like apices in the context. Basidiospores mostly oblong, double walls, (5.8-)6.4–7.3–7.8(-8.2) × (4.1-)4.6–6.1–6.7(-7.1) μm ( = 7.4 × 6.0 μm, n = 50) μm, with Q = 1.01–1.12, L = 7.32 µm, W = 6.98 µm (including myxosporium), (5.2-)5.7–6.5–7.1(-7.5) × (3.7-)4.3–5.5–6.1(-6.5) μm ( = 6.62 × 5.58 μm, n = 50) μm, with Q = 1.14–1.24, L = 6.56 µm, W = 5.48 µm (including myxosporium), overlaid by hyaline, dextrinoid, echinulate, inner wall echinulate, grayish-orange (5B4–5B5) in 5% KOH, and yellowish-brown (5E8) to light brown (6D8). Basidia not seen.
Habitat: Solitary on rotten wood of unknown tree species.
Specimens examined: THAILAND, Chiang Mai Province, Mae Taeng, Mushroom Research Centre, 19°07′200′ N, 98°41′44″ E, 770 m elev., 14 June 2019, P. E. Mortimer, MFLU 19-2222 and MFLU 19-2223.
Notes: Ganoderma philippii was introduced as
Fomes philippii by Bresadola and Hennings in 1891 [
126] and later transferred to
Ganoderma [
157].
Ganoderma philippii causes red root-rot disease, one of the most economically important diseases across a wide range of commercial perennial woody crops of tropical
Acacia species [
158].
Ganoderma philippii is distributed across Asia, from the Philippines in the north to southern Papua New Guinea [
111]. This species is characterized by its sessile, non-laccate pileus surface, with melanoid bands that form a layer distinct from the context, with a di-trimitic hyphal system with clamped connections overlaid by hyaline, echinulate, inner wall echinulate, sometimes turgid vesicular appendix basidiospores [
26]. In this study, our new record of
G. philippii collected from Thailand is described based on characteristics and phylogenetic analyses. Our collection agrees well with the description by Wang et al. [
33].
Ganoderma sichuanense S.H. Wu, Y. Cao, and Y.C. Dai. Fungal Diversity. 56(1): 54 (2012) (
Figure 30)
Facesoffungi number: FoF 06248
Description: Basidiomes annual or perennial, stipitate. Pileus 15 cm in length, up to 10 cm in width, and 2 cm thick. Pileus shell-like (involute from margin into the center), subflabellate or reniform to circular when seen from above, often with undefined concentric zones at center and extend to the margin, thick at center slightly soft at margin, and leathery when age when break. Pileus surface usually laccate, faded or week laccate when young, and strongly laccate when mature to age, shiny, silky, smooth, and soft when fresh, furrowed on the surface with sulcate to undulating, somewhat spathulate to uneven, incised, hard, and woody when old, and some occurred the brown (7D5) lined when older. Pileus color usually homogenous at base at the center with red (9A6–9A7, 9B7–9B8), brownish-red (9C6–9C8), and dark red (10C7–10C8), extended to the margin with reddish-yellow (4A7), but do not change the color when touched. Context up to 0.3–0.5 cm thick at base, abundant thick-walled, some thin-walled, with subsolid hyphae, bearing clamp connections, and occurred the dark brown (7F7–7F8) melanoid bands. Tube layers usually thin-walled, frequently branched with clamp connections, and hard and woody when mature, often brown (7D7). Stipe up to 3–8 cm in lenge, up to 1–3 cm in width, centrally stipitate, almost sub-cylindrical to cylindrical, often red (9A6) to brownish-red (9C6), and dark red (10C7) when mature. Margin obtuse from the center, some wavy, slippery when wet, softer, strong laccate edged, thin than the base and soft than the center, often reddish-yellow (4A7, 4B7–4B8), deep yellow (4A8), and orange (5A6–5A7) to golden yellow (5B7–5B8) when mature to old. Pore angular, subcircular to circular, 4–7 in number per mm, (40-)80–140(-155) μm. Pore surface white (11A1) to yellowish-white (3A2) when young to mature, turning yellow (3B8) to olive-yellow (3C7–3C8) when dried, as well as becoming discolored when touched, turns light brown (7D5), brown (7D7–7D8) to dark brown (7F6–7F8) when scratched or bruised.
Hyphal structure: Hyphal system trimitic, with bearing clamp connections, hyaline, with walls varying in thickness with simple septa, composed of narrow and sparingly branched; generative hyphae (1.3-)1.6–1.8–2.2(-2.4) μm broad (n = 30), thin-walled, hyaline, branched, with clamp connections; skeletal hyphae (1.6-)2.2–3.5–4.2(-4.9) μm broad (n = 30), walls varying in thickness, sometimes subsolid; binding hyphae (1.2-)1.5–1.7–2.0(-2.3) μm broad (n = 30), usually thick-walled with narrow to subsolid, with pale orange (5A3) to light orange (5A4) of thin-walled, and orange (6B7) to brownish-orange (6C8) of thick-walled in Melzer’s reagent, and occurred the melanoid bands. Pileipellis a hymeniderm, brown (7D7) with clavate-like cells, dextrinoid. Basidiospores mostly ellipsoid or oblong ellipsoid, truncate at maturity, with double walls, (8.0-)9.4–11.3–12.2(-13.3) × (4.9-)5.4–6.5–7.1(-7.4) μm, ( = 11.2 × 6.6 μm, n = 50) μm, with Q = 1.64–1.51, L = 11.24 µm, W = 6.69 µm (including myxosporium), (7.8-)8.1–8.6–9.9(-11.7) × (4.9-)5.2–5.9–6.7(-7.4) μm (Qm = 1.5, Q = 1.0–2.0, excluding outer myxosporium) (n = 50), ( = 8.7 × 5.8 μm, n = 50) μm, with Q = 1.46–1.53, L = 8.72 µm, W = 5.84 µm (excluding outer myxosporium), overlaid by hyaline, exosporium (outer wall) smooth and hyaline, endosporium (inner wall) coarse echinulate, with turgid vesicular appendix, light brown (7D7) to brown (7D8) in KOH, and reddish-brown (8E6) to dark brown (8F4) in Melzer’s reagent. Basidia 12–17 × 6–9 μm, with four sterigmata.
Habitat: Occasionally occurring on the stump of decaying Pterocarpus sp., living tree.
Specimens examined: THAILAND, Prachuap Khiri Khan Province, 12°08′52″ N, 99°45′41″ E, 491 m elev., 25 June 2018, T. Luangharn, MFLU 19-2164.
Ganoderma sinense J.D. Zhao, L.W. Hsu, and X.Q. Zhang, Acta Mycologica Sinica. 19: 272 (1979) (
Figure 31)
Facesoffungi number: FoF 06253
Description: Basidiomes annual, stipitate, subdimidiate. Pileus 4–10 cm in length, 6–12 cm in width, 0.3–1.3 cm thick at base. Pileus stipitate, subdimidiate to dimidiate, flabelliform, spathulate, umbonate, concentrically sulcate zone, radial from center extending to the margin, tough when broken, often thick at center, slightly soft at margin, light in weight when dried, and non-woody. Pileus surface laccate, convex, some radial furrowed, imbricate, incised, glossy, shiny, spathulate, shallow sulcate when fresh, umbonate or uneven, strongly laccate and glossy when mature, and weakly laccate where the new hyphae are in active development (margin), usually smooth layers at center from young to old age, irregularly ruptured crust overlying the context, and leathery when broken in old age. Pileus color usually homogenous and brownish-red (8C7–8C8) to reddish-brown (8D7–8D8) at center toward stipe and extending brownish-red (9C8) from the center slight to the margin when mature, usually reddish-brown (8E5–8E8) upper margin surface when old. Context up to 0.3–0.8 cm thick near the stipe, dried, upper layers brownish-orange (6C8) when fresh, lower layers grayish-orange (5B5), dark brown (8F7) when dried, soft and fibrous, covered with thin crust, some present woody, trimitic hyphae, hyaline, walls varying in thickness with simple septa, with branches. Tube 0.3–0.9 cm in length, brown (7D8). Stipe 3–12 cm in length, sub-cylindrical to cylindrical, almost stipitate, with broad and thick at base, irregularly ruptured crust overlying, usually strongly laccate, brown (7D8) to dark brown (8F8) when mature, usually dark brown (8F8) when old. Margin soft, some wavy, laccate when mature, weakly laccate when old, brownish-orange (6D8) from mature to old. Pore 4–6 in number per mm, subcircular to circular. Pore surface white (11A1) when present, to yellowish-white (2A2) when mature, turns brownish (6E7) to dark brown (6F7) when scratched or bruised.
Hyphal structure: Hyphal system trimitic, with clamp connections, usually light orange (5A5) to orange (5A7), golden yellow (5B7–5B8), sometimes brownish-red (8C7) in KOH; generative hyphae 1.2–2.1 µm broad (n = 30), hyaline, thin-walled, with clamp connections; skeletal hyphae 3.2–5.4 µm broad (n = 30), usually hyaline, thick-walled, unbranched, and solid; binding hyphae 3.3–5.7 µm width (n = 30), walls varying in thickness, with many branches, hymenial with sword-like apices in the context. Pileipellis a hymeniderm, grayish-orange (5B5) to brown (6E8), clavate-like cells, dextrinoid. Basidiospores mostly ellipsoid to broadly ellipsoid, double walls, (9.8-)10.4–11.7–12.5(-13.4) × (7.3-)7.7–8.9–10.0(-10.6) μm ( = 11.8 × 9.0 μm, n = 50) μm, with Q = 1.29–1.34, L = 11.82 µm, W = 9.02 µm (including myxosporium), (8.9-)9.8–10.4–12.0(-12.8) × (6.2-)6.8–7.2–7.9(-8.3) μm ( = 10.6 × 7.3 μm, n = 50) μm, with Q = 1.41–1.47, L = 10.47 µm, W = 7.25 µm (excluding outer myxosporium), overlaid by hyaline, apically and shortly echinulate, truncate, some turgid vesicular appendix, inner wall echinulate, orange (5A7), deep (5A8, 5B8), orange (6B8), brownish-orange (6B8), outer wall usually brownish-red (8C7–8C8) in 5% KOH.
Habitat: Solitary, on the living tree of Dendrocalamus strictus and Dipterocarpus sp.
Specimens examined: THAILAND, Nakhon Phanom Province, 22 December 2018, T. Luangharn, MFLU 19-2172.
Ganoderma subresinosum (Murrill) C.J. Humphrey, Mycologia. 30 (3): 332 (1938) (
Figure 32)
≡ Fomes subresinosus Murrill, Bulletin of the Torrey Botanical Club. 35: 410 (1908)
≡ Trachyderma subresinosum (Murrill) Imazeki, Bulletin of the Government Forest Experimental Station Meguro. 57: 119 (1952)
≡ Magoderna subresinosum (Murrill) Steyaert, Persoonia. 7(1): 112 (1973)
≡ Amauroderma subresinosum (Murrill) Corner, Beihefte zur Nova Hedwigia. 75: 93 (1983)
= Polyporus mamelliporus Beeli, Bulletin de la Société Royale de Botanique de Belgique. 62(1): 62 (1929)
Facesoffungi number: FoF 06259
Description: Basidiomes annual, subdimidiate, sessile. Pileus 6–13 cm in length, 4–10 cm in width, and 0.6–2 cm thick. Pileus stipitate, flabelliform, spathulate, umbonate, subdimidiate to dimidiate, single, concentrically sulcate zone, radial from center extending to the margin, tough when broken, often thicker at center, slightly soft at margin, and light in weight when dried. Pileus surface convex, glossy, shiny, usually frequently furrowed, shallow sulcate, mostly rugulose, spathulate, umbonate to uneven, laccate when mature, strongly laccate when old, concentrically sulcate with irregularly ruptured crust overlying the context, hard from mature to old, and woody or corky texture when old. Pileus color usually dark brown (9F7–9F8) from center to the margin. Context up to 0.3–1 cm thick near the base, dry, upper layers grayish-orange (5B6) when fresh, lower layers grayish-orange (5B5), dark brown (8F7) when dried, covered with crust, woody when dried, trimitic hyphae, hyaline, walls varying in thickness, with simple septa, with branches. Tube 0.4–1.2 cm in length, brown (7D8). Stipe almost sessile, blunt, broadly attached, and thick at base, irregularly ruptured crust overlying, and usually strongly laccate with brown dark brown (9F7) from mature to old. Margin strongly laccate and dark brown (9F7) when mature to old. Pore 4–5 in number per mm, subcircular to circular. Pore surface pale yellow (4A3) to pale orange (5A3), turns dark brown (6F7) when scratched or bruised.
Hyphal structure: Hyphal system di-trimitic, with clamp connections, usually light yellow (3A5) to grayish-yellow (3B5), pale yellow (4A5), light orange (5A5) to orange (5B8) in KOH; generative hyphae 1.0–2.1 µm broad (n = 30), thin-walled, hyaline, with clamp connections; skeletal hyphae 2.8–5.2 µm broad (n = 30), usually unbranched or few branches, and thick-walled; binding hyphae 2.5–5.0 µm width (n = 30), walls varying in thickness, many branches, some hymenial with sword-like apices in the context. Pileipellis a hymeniderm, light orange (5A5), clavate-like cells, dextrinoid. Basidiospores mostly ellipsoid to broadly ellipsoid, double walls, (11.6-)12.1–13.5–14.5(-15.8) × (8.1-)8.9–11.3–11.9(-12.5) μm ( = 13.2 × 11.4 μm, n = 50) μm, with Q = 1.12–1.19, L = 13.24 µm, W = 11.44 µm (including myxosporium), (10.2-)10.9–12.8–13.7(-14.6) × (6.8-)7.9–8.9–9.6(-10.2) μm ( = 12.6 × 8.8 μm, n = 50) μm, with Q = 1.41–1.46, L = 12.63 µm, W = 8.84 µm (excluding outer myxosporium), overlaid by hyaline, apically and echinulate, truncate, inner wall usually orange (6A7–6A8) to reddish-golden (6C7) in KOH, brownish-orange (5C5–5C6) in Melzer’s reagent, outer wall usually reddish-orange (7A8, 7B8) in 5% KOH and brown (6D7–6D8) in Melzer’s reagent.
Habitat: Solitary on the decaying hardwood of Peltophorum pterocarpum and Castanopsis sp.
Specimens examined: THAILAND, Chiang Rai Province, 20°15′03″ N, 100°14′17″ E, 732 m elev., 9 October 2017, T. Luangharn, MFLU 17-1912.
Notes: Ganoderma subresinosum was introduced as
Fomes subresinosus with the specimen from the Philippines [
43]. Next, Humphrey [
159] verified this fungus species to the genus
Ganoderma, and then Imazeki [
160] included this species in the genus
Trachyderma.
Ganoderma subresinosum is a species that is distributed worldwide, known from the Philippines to other Asian countries, and distributed across Eastern and Central Africa [
30,
47,
100,
110]. This fungus is distinctive in form with its laccate pileus, sessile, dark brown pileus surface, and concentrically sulcate with irregularly ruptured crust overlying the context, with ellipsoid to elongate basidiospores.
Ganoderma subresinosum was regarded as a synonym of
Trachyderma tsunodae Imazeki [
160],
Magoderna subresinosus [
110], and
Amauroderma subresinosum [
113]. In China, this fungus was reported by Chinese researchers as
Fomes subresinosum [
132,
161,
162] and verified to
A. subresinosum by Zhao [
61] and Zhao and Zhang [
13]. Recently, several studies have suggested that those three synonymous fungal species are different from
G. subresinosum based on emergent morphological and molecular data [
163,
164]. However, this fungus name is in the Index Fungorum as
G. subresinosum. So, in this study, we present the first record of
G. subresinosum from Thailand.
Ganoderma thailandicum T. Luangharn, P.E. Mortimer, S.C. Karunarathna, and J.C. Xu, MycoKeys 59: 55 (2019)
Facesoffungi number: FoF 06129
Index Fungorum number: IF 556535
For characteristics, see Luangharn et al. [
31].
Notes: Ganoderma thailandicum is characterized by its laccate, deep magenta close to stipe, brownish-red at center, and light yellow around active development toward the margin on pileal surface, white pore surface, brownish-red context, and absence of melanoid band.
Ganoderma tropicum (Jungh.) Bres., Annales Mycologici 8(6): 586 (1910)
≡ Polyporus tropicus Jungh., Praemissa in floram cryptogamicam Javae insulae: 63 (1838)
≡ Fomes tropicus (Jungh.) Cooke, Grevillea. 14 (69): 19 (1885)
≡ Scindalma tropicum (Jungh.) Kuntze, Revisio generum plantarum. 3 (2): 519 (1898)
Facesoffungi number: FoF 05068
For characteristics, see Luangharn et al. [
75].
Notes: Ganoderma tropicum was introduced as
Polyporus tropicus by Junghuhn [
165] with the specimen from Indonesia. Phylogenetic analysis has been well resolved with the aid of molecular data [
5,
129] and transferred this species to
Ganoderma [
166], where it is considered a member of the
G. lucidum species complex [
76,
167]. The fungus is characterized by its distinctive reddish-brown pileal surface, dark brown context near the tubes, dense context layer, thick near the base, with strongly echinulate basidiospores. Its distribution is highly variable worldwide, scattered across tropical Asian regions, mainland China [
4,
5,
30], South America [
137], and Taiwan, PRC [
33], and it causes white root and butt rot on several tree species [
8]. Furthermore, there are some similarities between
G. tropicum and other
Ganoderma species. According to Cao et al. [
4], among the Chinese
Ganoderma species,
G. flexipes,
G. multipileum,
G. sichuanense, and
G. tsugae are similar to
G. tropicum, having a reddish-brown pileus surface, dark brown context, ellipsoid basidiospores, strongly echinulate basidiospores, and irregular cuticle cells. Our new record of
G. tropicum from Northern Thailand was described based on characteristics together with phylogenetic analyses, the details of which are shown in Luangharn et al. [
75].
Taxonomy of Ganoderma from Vietnam
Ganoderma hochiminhense Karunarathna, Mortimer, & Luangharn, sp. nov. (
Figure 33)
Facesoffungi number: FoF 06334
Index Fungorum number: IF 556794
Diagnosis: Ganoderma hochiminhense is characterized by its strongly laccate appearance, with a reddish-brown color near the stipe, a deep orange to brownish-yellow at the center, a white that indicates active development on the margin of the pileal surface, a yellowish-white pore surface when fresh, an orange, deep orange to reddish-orange context, and absence of melanoid bands.
Holotype: Vietnam, Hochiminh City, on Areca sp. (as described by the seller), 12 June 2019, LT2019 Gano 305 (MFLU 19-2224) and LT2019 Gano 306 (MFLU 19-2225).
Etymology: The specific epithet “hochiminhense” refers to the place in Vietnam from where the holotype specimen was collected.
Description: Basidiomes sessile, orbicular. Pileus up to 0.8–2.2 cm in length, 0.5–1.4 cm width, and 1.3 cm thick at base. Pileus orbicular, undulated, tuberculate, sulcate, single or fused at the base, broadly attached, hard when dried. Pileus surface distinctively laccate when young, strongly laccate when mature or dried. Pileus color reddish-brown from the base, light to deep orange, golden yellow, brownish- to reddish-yellow at the center, with deep yellow to orange yellow at the margin of the upper surface, generally white where new hyphae are in active development, glossy, shiny, smooth, spathulate, sulcate when fresh, with a thin crust overlying the pileus, which is thicker at the base than at the margin, light weight when dried. Hymenophore mostly orange to brownish-orange in KOH, up to 0.8 cm thick, with a dense but not fully homogenous context layer, bearing distinct layers of concentric growth zones at the center that extend to the margins, thick near the base, non-corky or woody texture when dried, bearing a simple septum at base, tough to break when dried, melanoid bands absent. Context reddish-orange in KOH; generative hyphae up to 2.72–3.82 μm ( = 3.54, n = 50) in diam, almost colorless, thin-walled, some expanded at the apex, unbranched, with clamp connections; skeletal hyphae-dominant, up to 3.56–7.47 μm ( = 5.85, n = 50), orange to brown, thick-walled, unbranched, without clamp connections; binding hyphae 3.23–5.96 µm ( = 4.32, n = 50), almost colorless, walls varying in thickness, with some narrow lumen to subsolid, frequently branched, tortuous, and interwoven at the distal end. Tubes hard, brown, up to 0.2–0.5 cm long. Tubes layers generative hyphae 2.21–3.03 µm in diam, pale brown to brown, thin-walled, some thick-walled, with clamp connections, unbranched; skeletal hyphae 3.23–6.15 µm in diam, distinctly brown, thick-walled, some narrow lumen to subsolid, frequently branched; binding hyphae 2.28–4.86 µm in diam, brownish-yellow, thick-walled to solid, and frequently branched. Stipe short stipe, laccate when developing to maturity, and strongly laccate from maturity to old age. Margin white when present to maturity, light brown to brown between the young to mature stages, turning light brown when dry, and silky, soft, and slippery to the touch between youth and maturity, usually bruising when touched and tough to break. Pore angular to round, 4–6 in number per mm, with dissepiments slightly thick to thick. Pore surface white when fresh, yellowish-white when dried, turns brown to dark brown when touched.
Hyphal structure: Hyphal system trimitic, with tissues yellow, light orange to orange and brown in KOH; generative hyphae up to 2.70–3.25 μm ( = 3.09, n = 50) in diam, almost colorless, thin-walled, unbranched, with clamp connections, some slightly swollen at the distal end; skeletal hyphae-dominant, up to 3.49–7.67 μm broad ( = 5.98, n = 50), pale brown to distinctly brown, thick-walled, unbranched, without clamp connections, occasionally with narrow lumen, some subsolid, interwoven; binding hyphae 3.12–5.87 µm broad ( = 4.28, n = 50), grayish-orange, walls varying in thickness, frequently branched, tortuous, and interwoven in the distal end. Basidia clavate, 4-sterigmatic, 10.2–14.3 × 8.6–12.5 µm, yellowish to pale brown in KOH. Basidiospores ellipsoid, sometimes broadly ellipsoid or almond-shaped at maturity, reddish-yellowish, light brown to brownish-orange, slightly truncate, double walls, exospore smooth, endospore with coarse echinulate, (6.8)8.5–10.4(11.8) × (5.8)6.9–9.3(10.2) μm ( = 9.4 × 8.8 μm, n = 50), with Q = 1.02–1.14, L = 9.38 µm, W = 8.82 µm (including myxosporium), (6.6)7.9–9.4(10.9) × (5.4)6.3–8.5(9.3) μm ( = 9.0 × 7.7 μm, n = 50) μm, with Q = 1.08–1.21, L = 8.98 µm, W = 7.73 µm (excluding outer myxosporium), strongly echinulate, cuticle cells irregular. Culture characteristics white active mycelium on PDA, reaching 8 cm diam after 14 days at 25 °C.
Habitat: solitary, on Areca spp. stumps.
Specimens examined: VIETNAM, Hochiminh City local market, 8°54′32″ N, 98°31′09″ E, 427 m elev., 12 June 2019, S.C. Karunarathna, LT2019 Gano 305 (MFLU 19-2224, holotype) and LT2019 Gano 306 (MFLU 19-2225, paratype).


 ,
, 


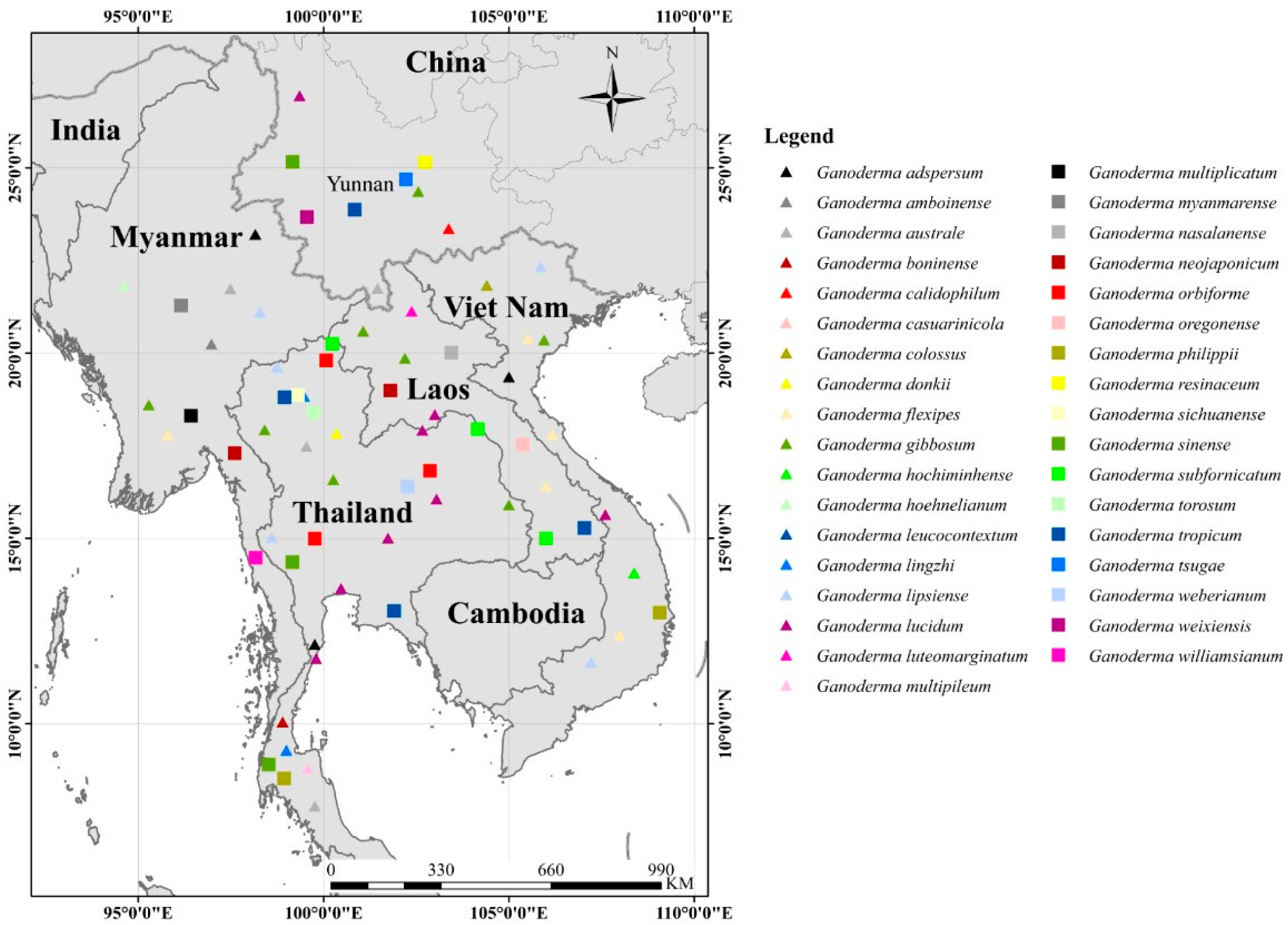{kind=link}
{kind=link}
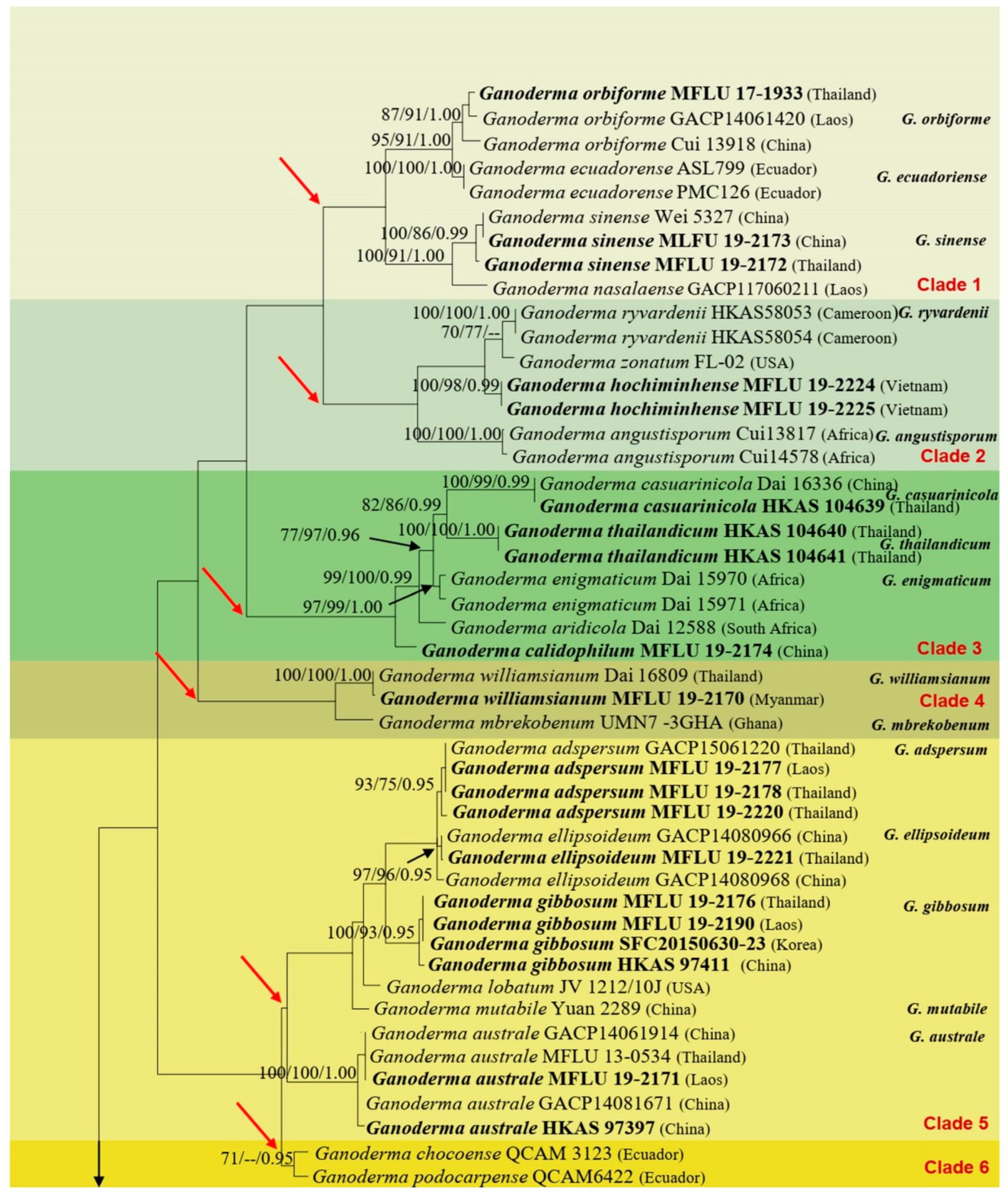{kind=link}
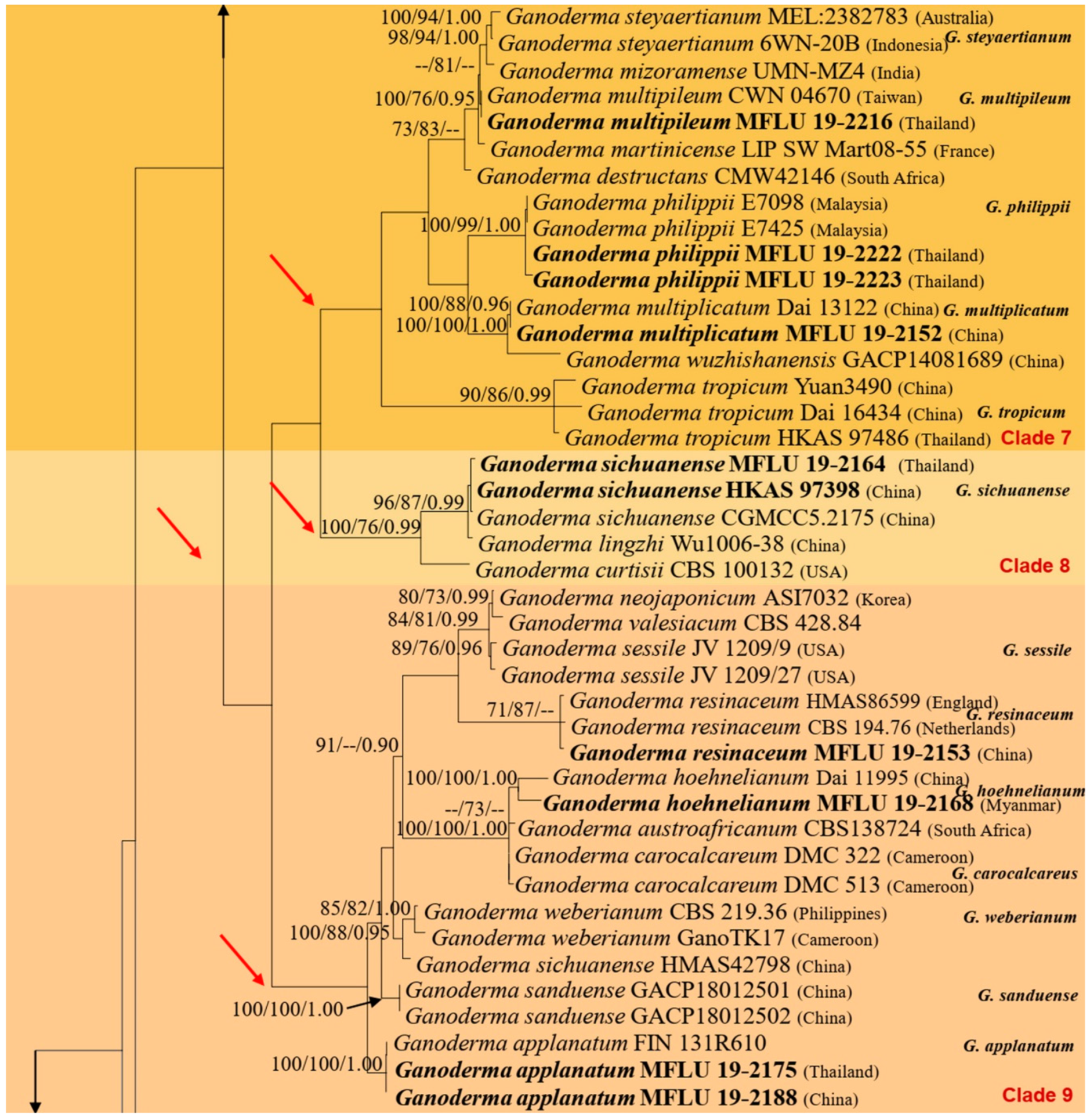{kind=link}
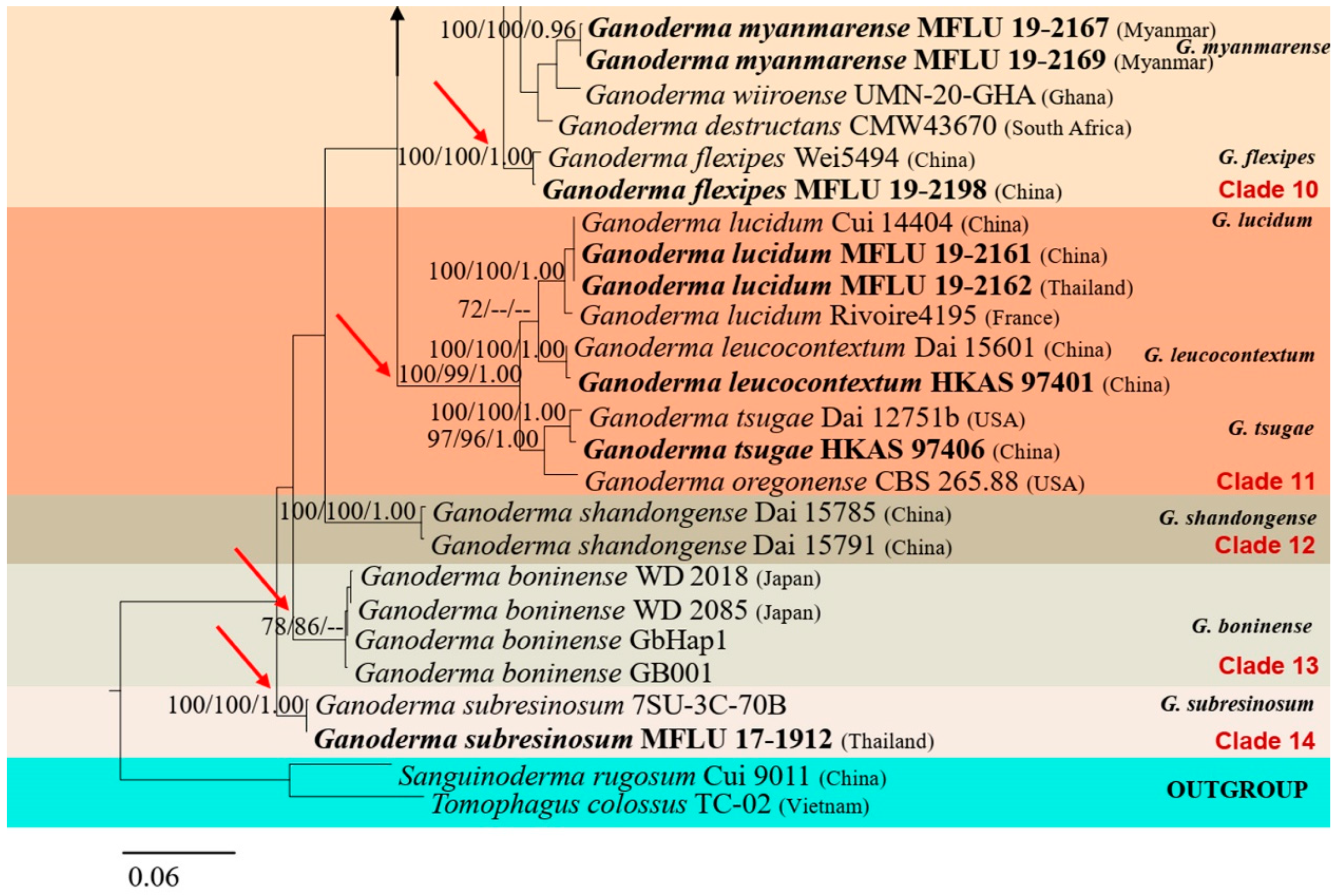{kind=link}
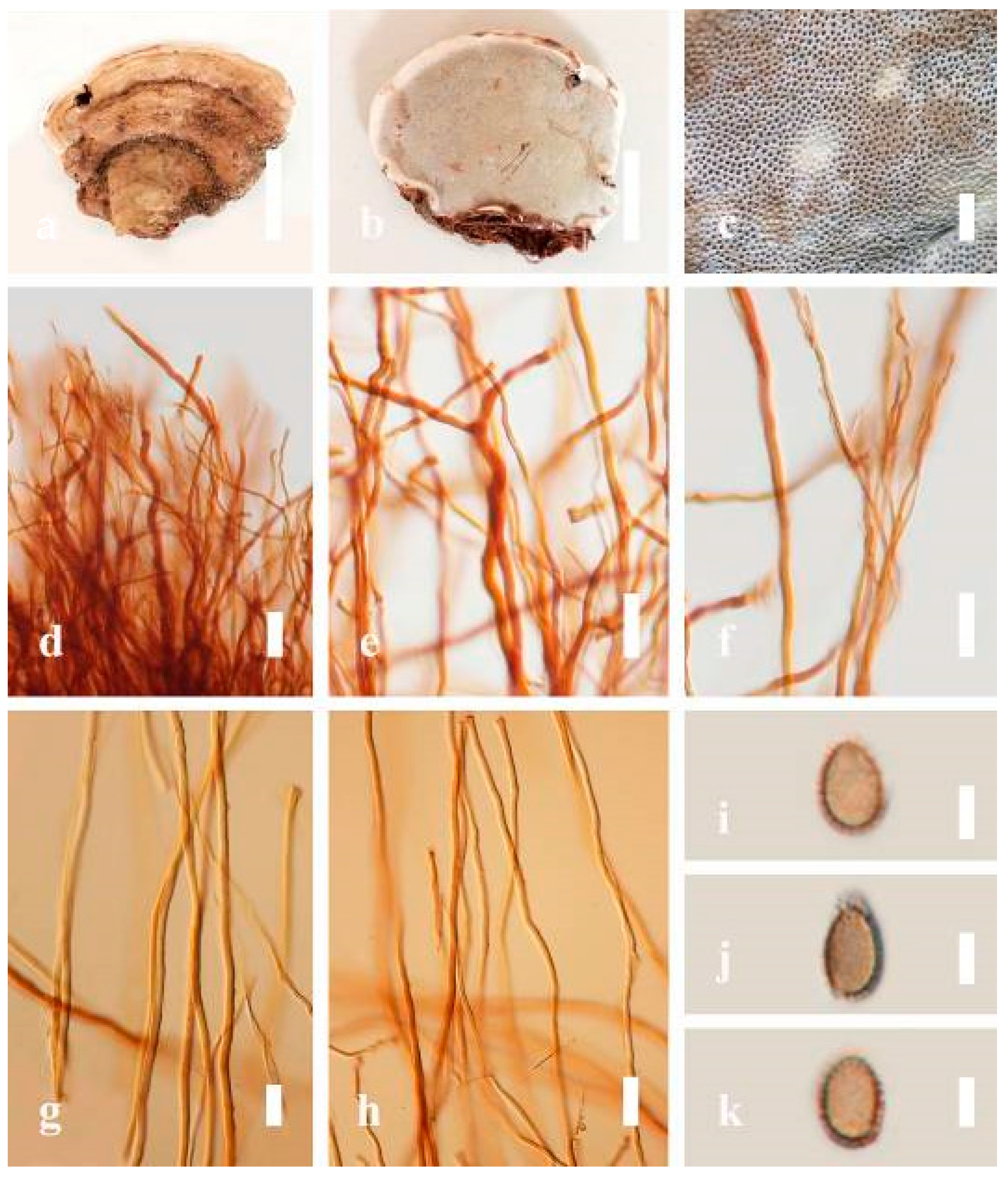{kind=link}
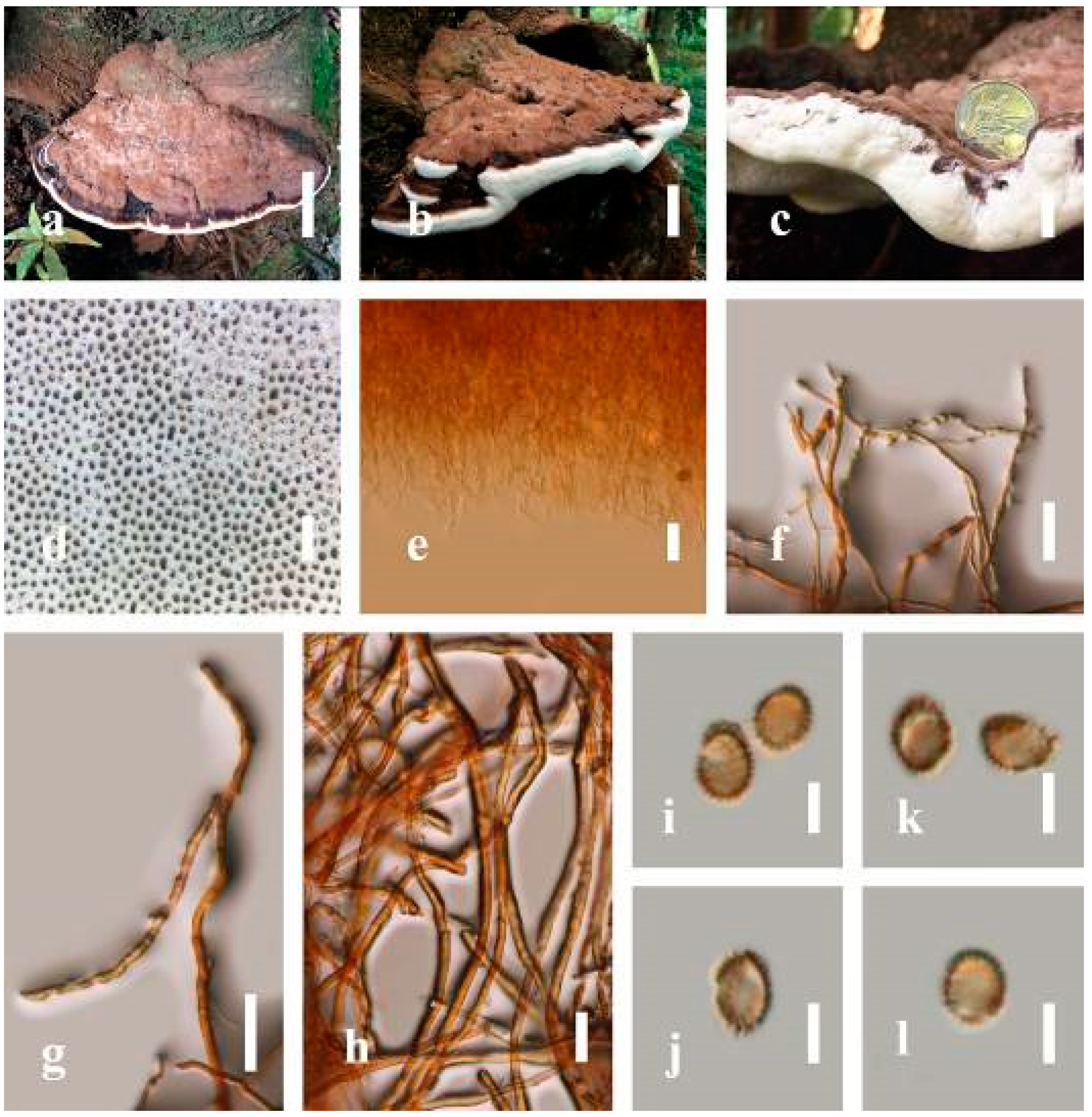{kind=link}
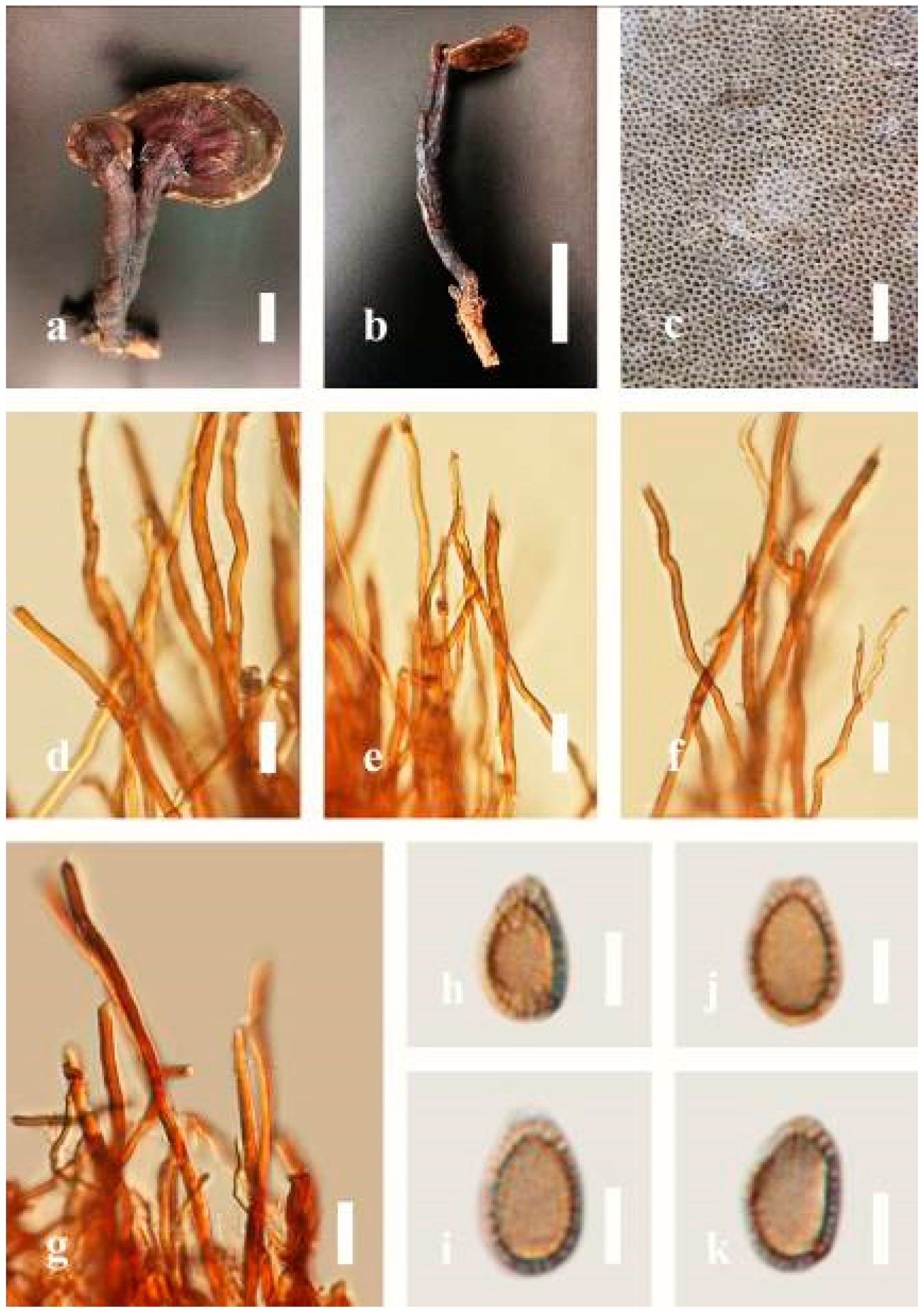{kind=link}
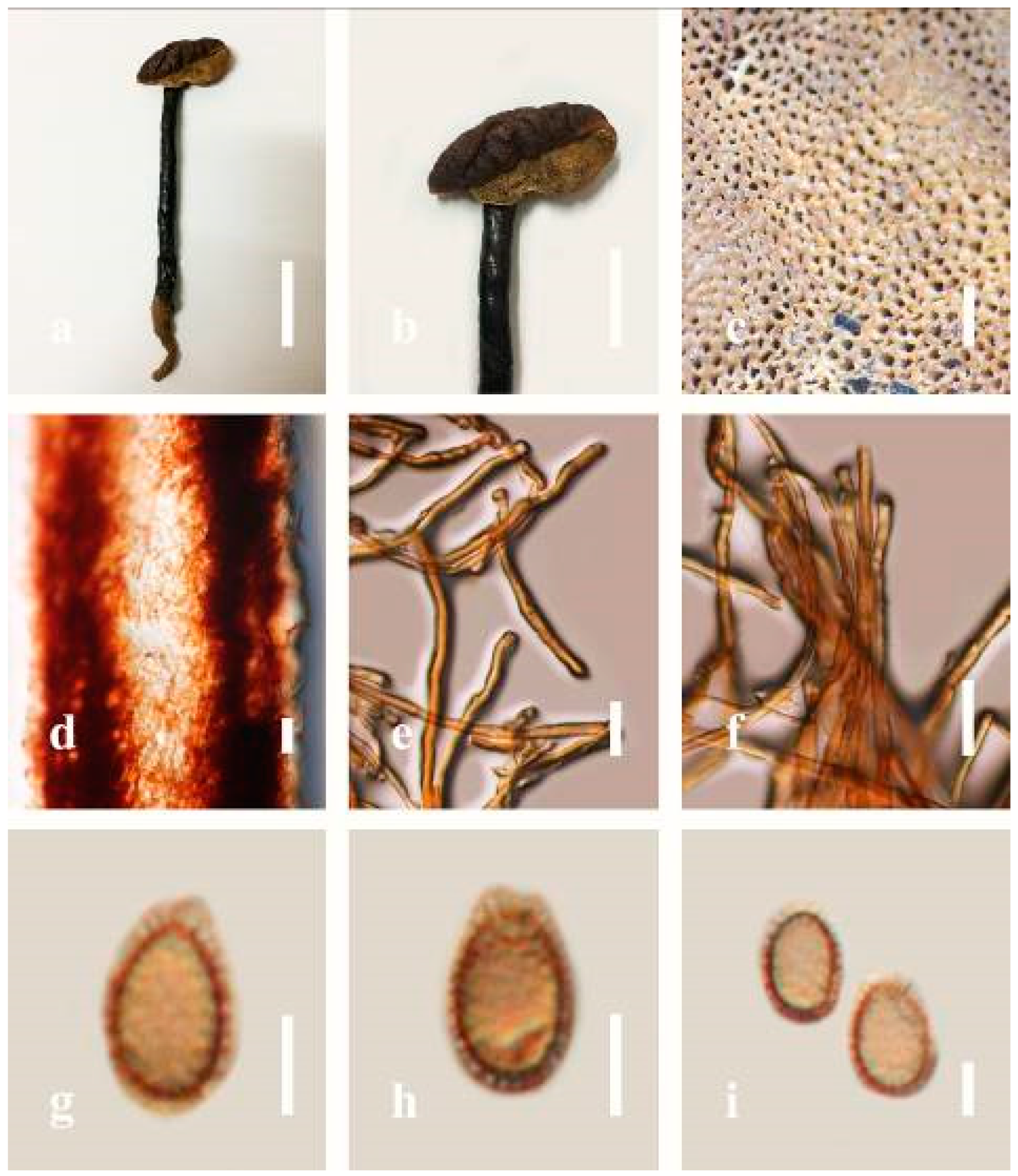{kind=link}
{kind=link}
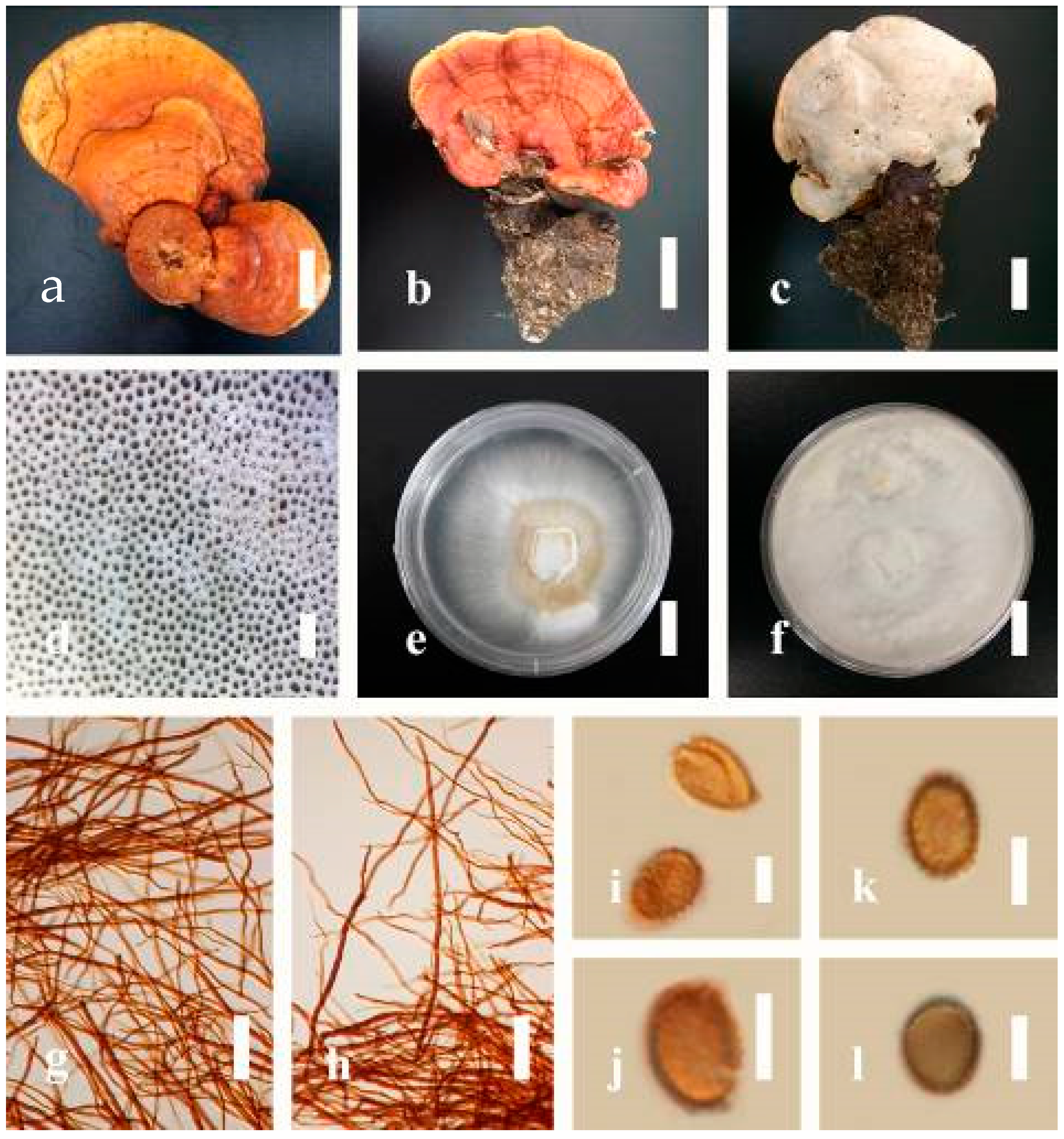{kind=link}
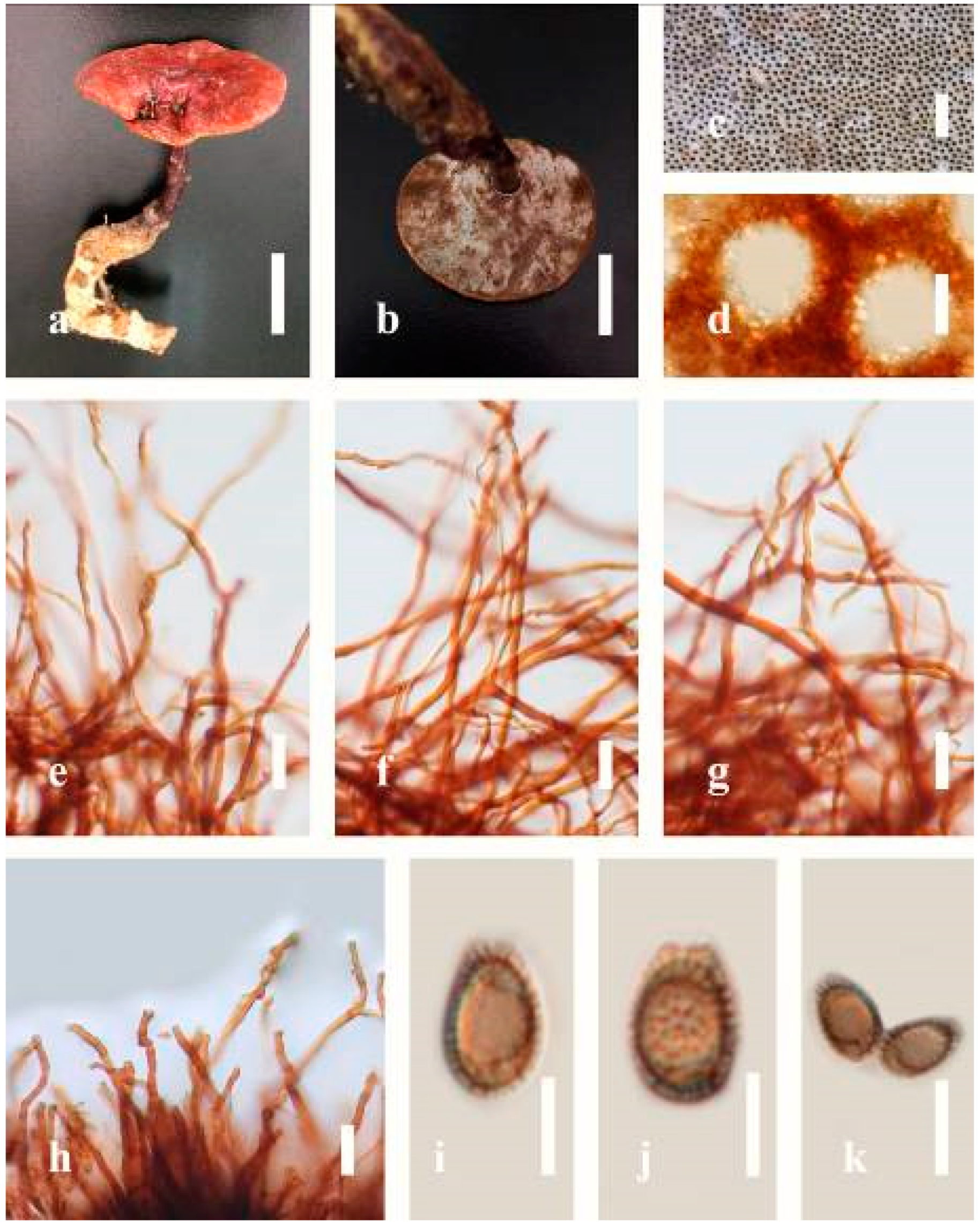{kind=link}
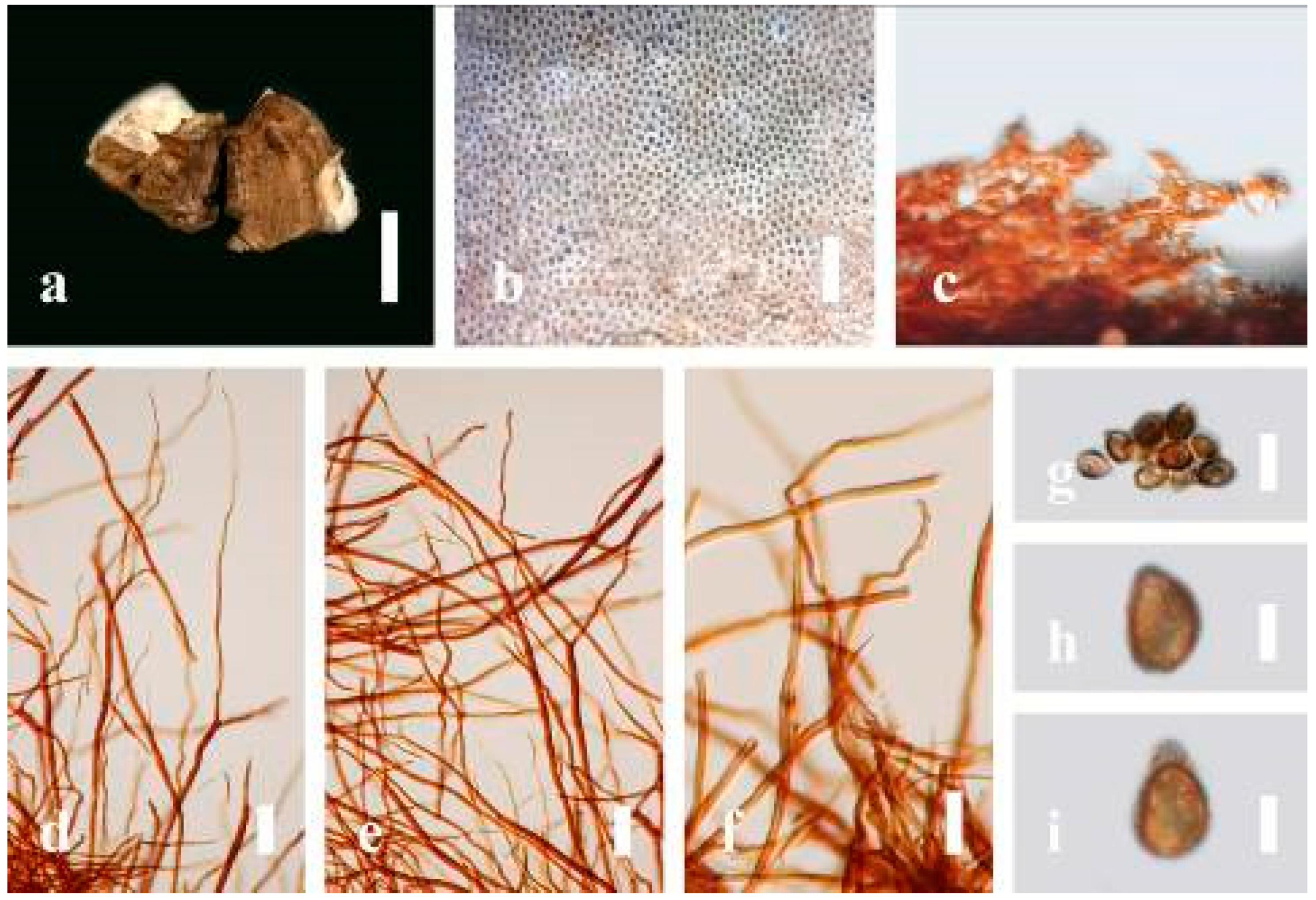{kind=link}
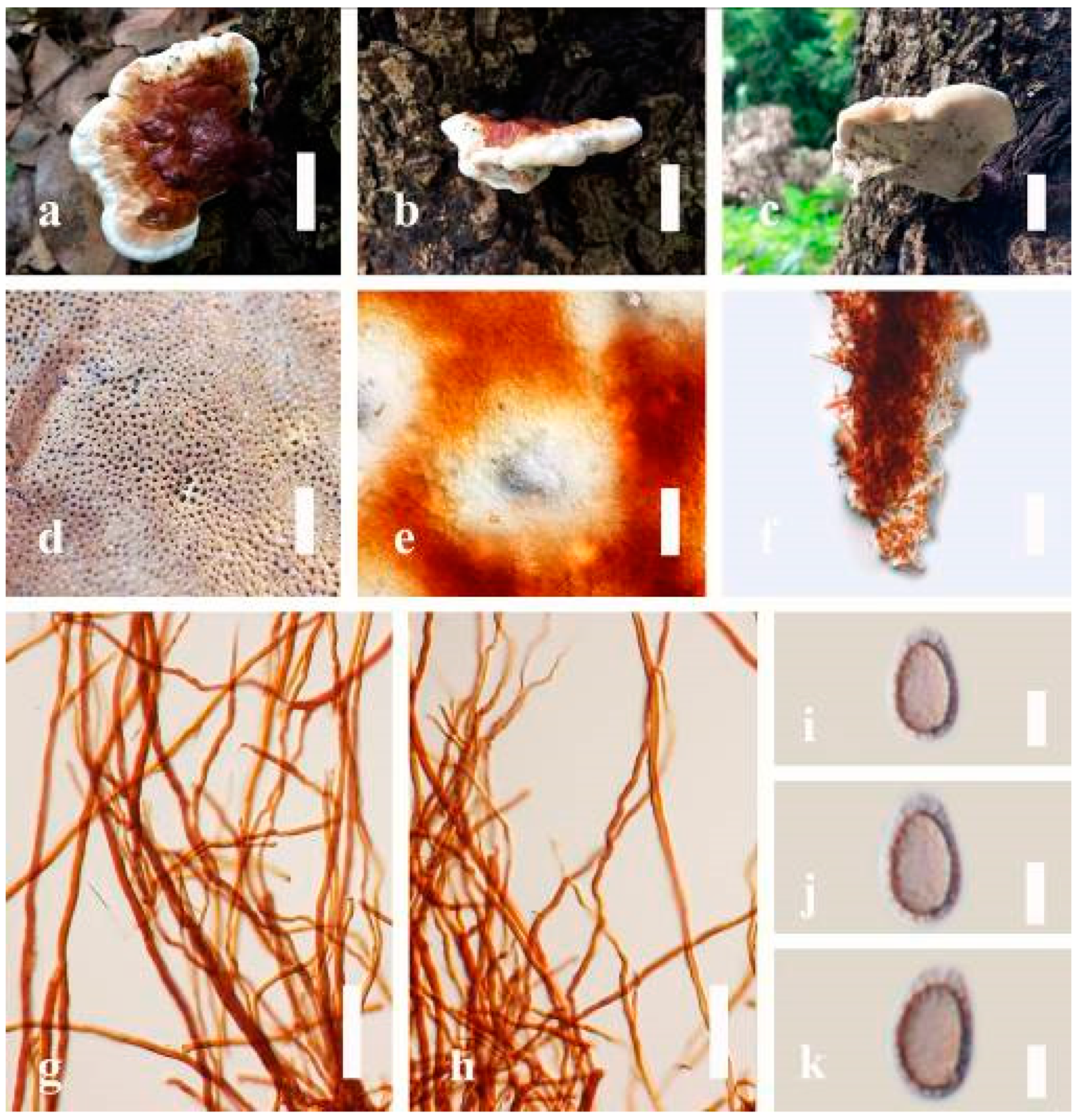{kind=link}
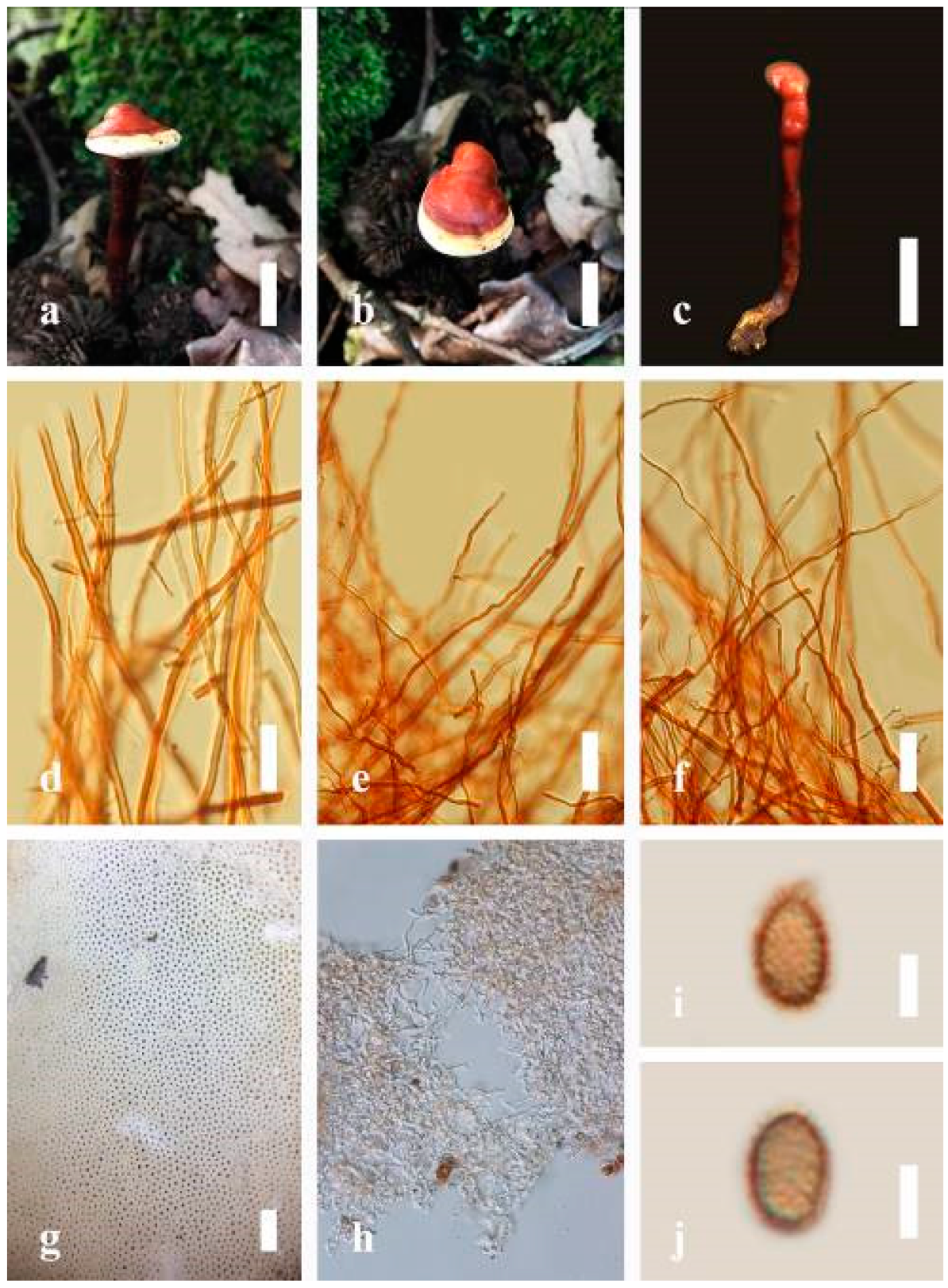{kind=link}
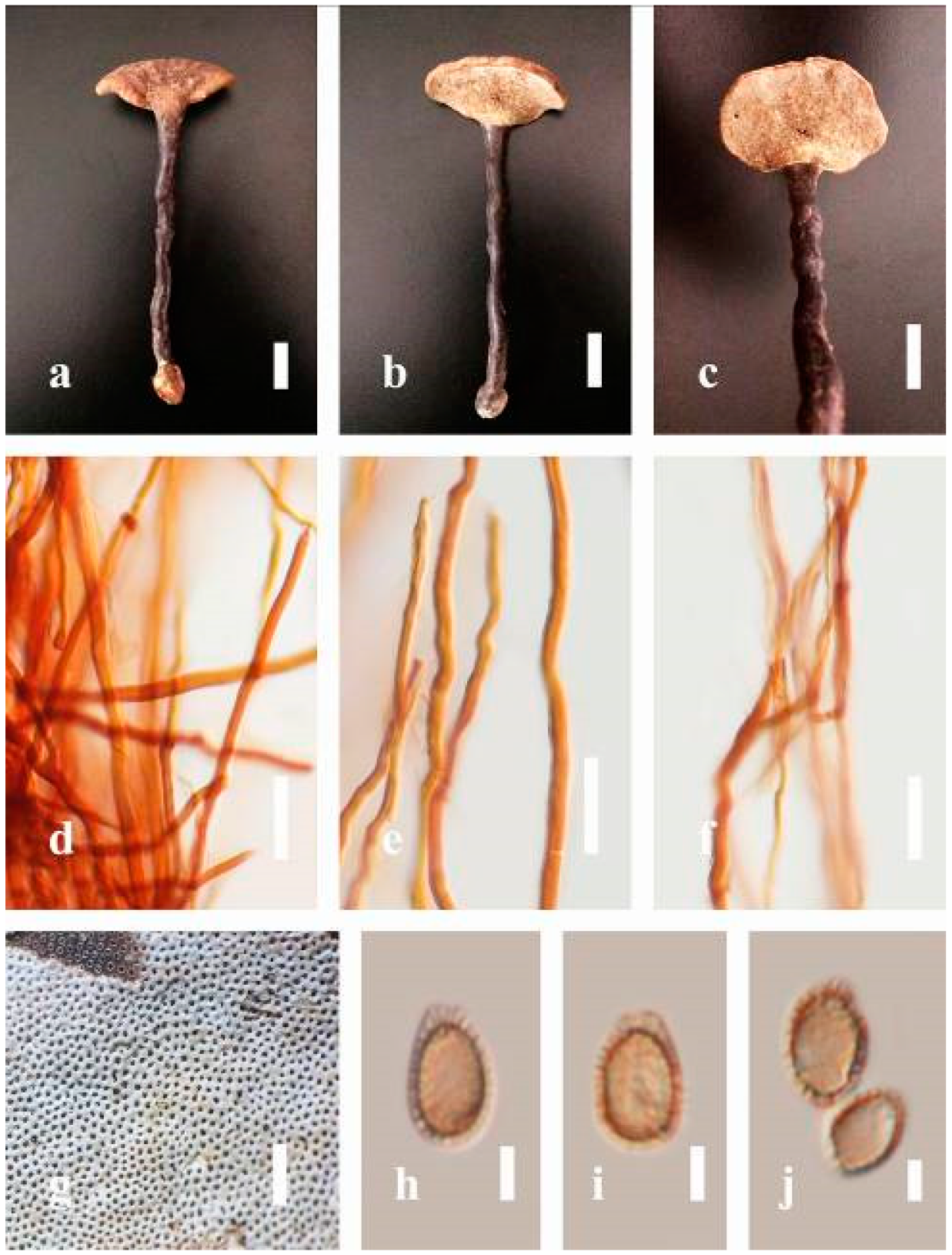{kind=link}
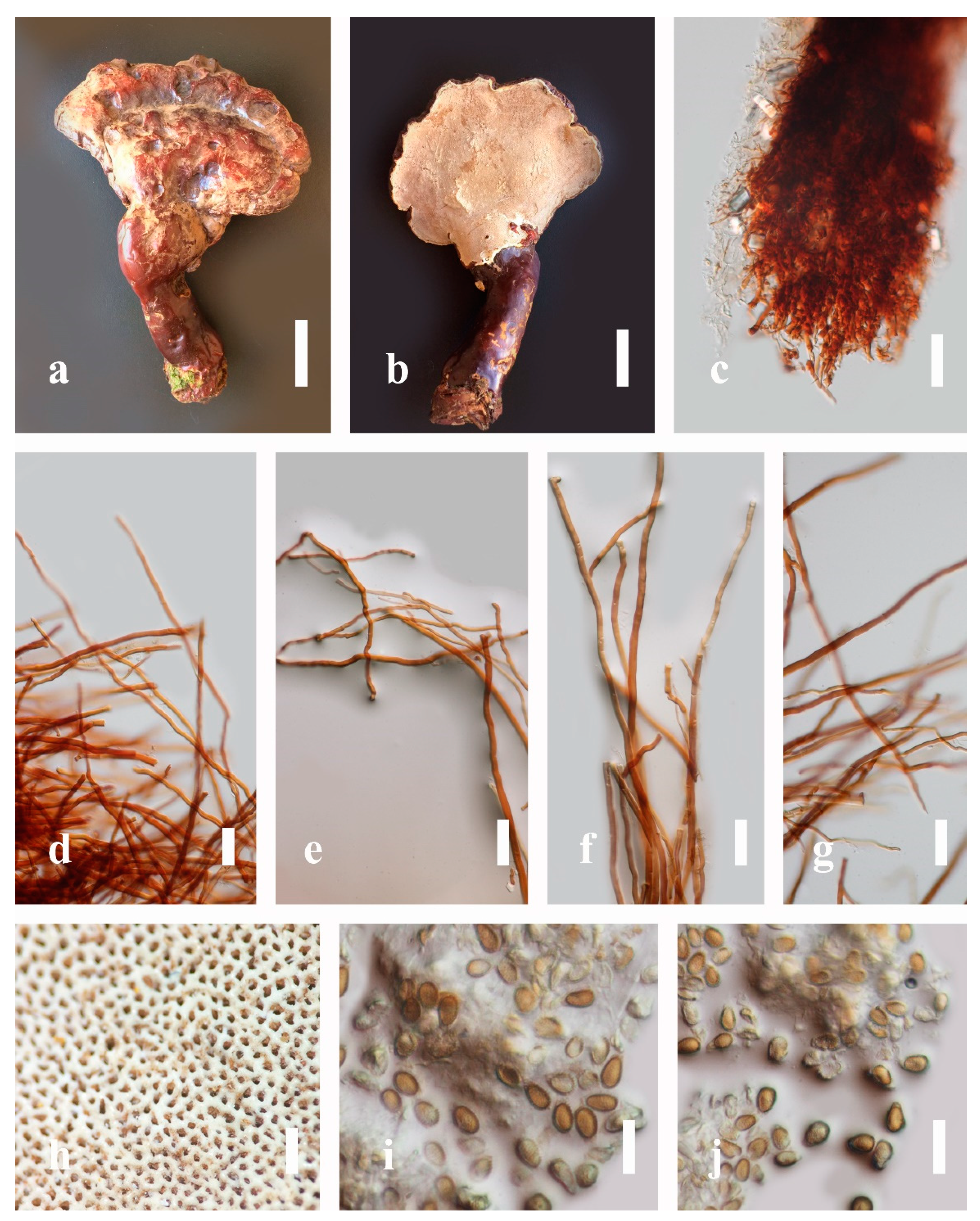{kind=link}
{kind=link}
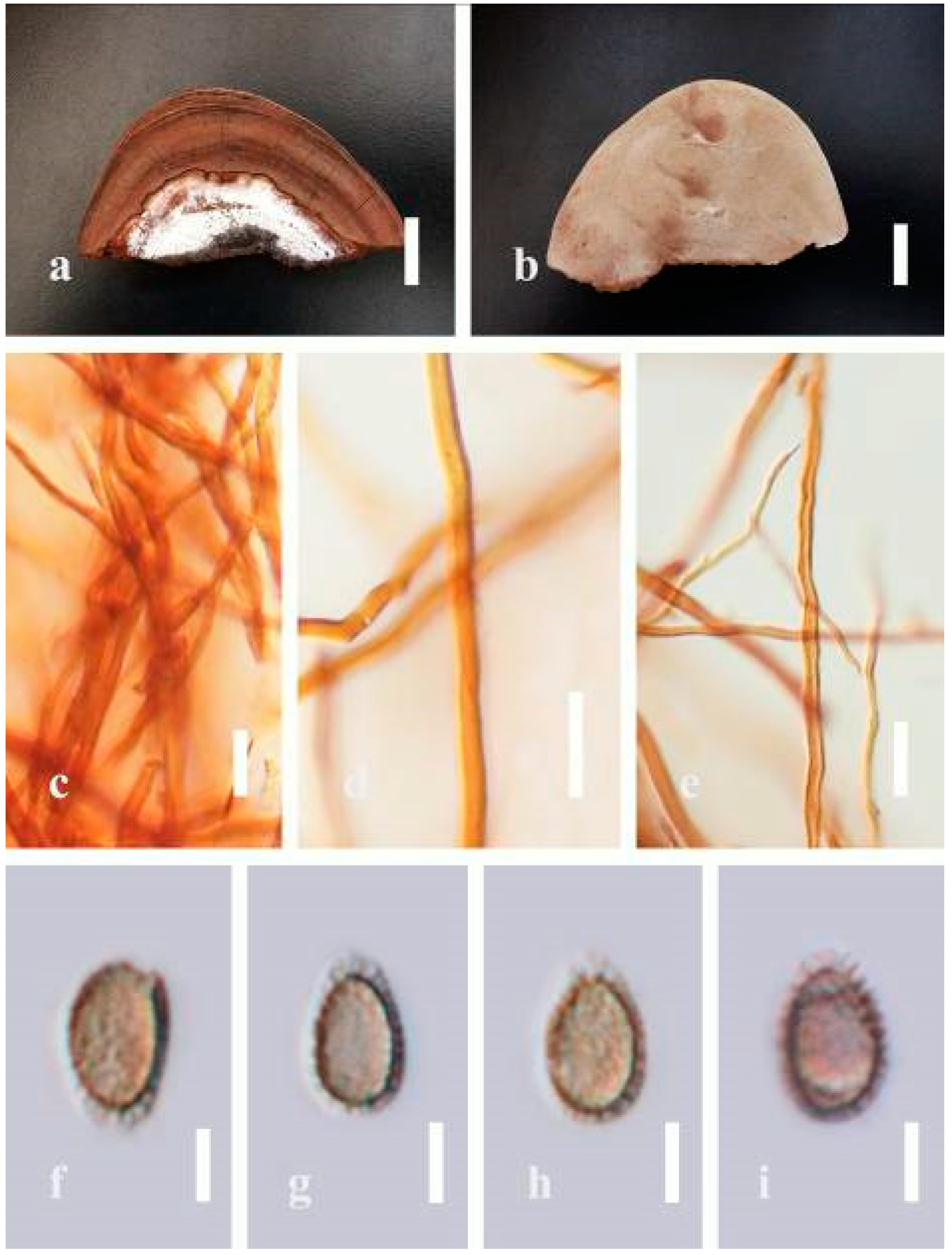{kind=link}
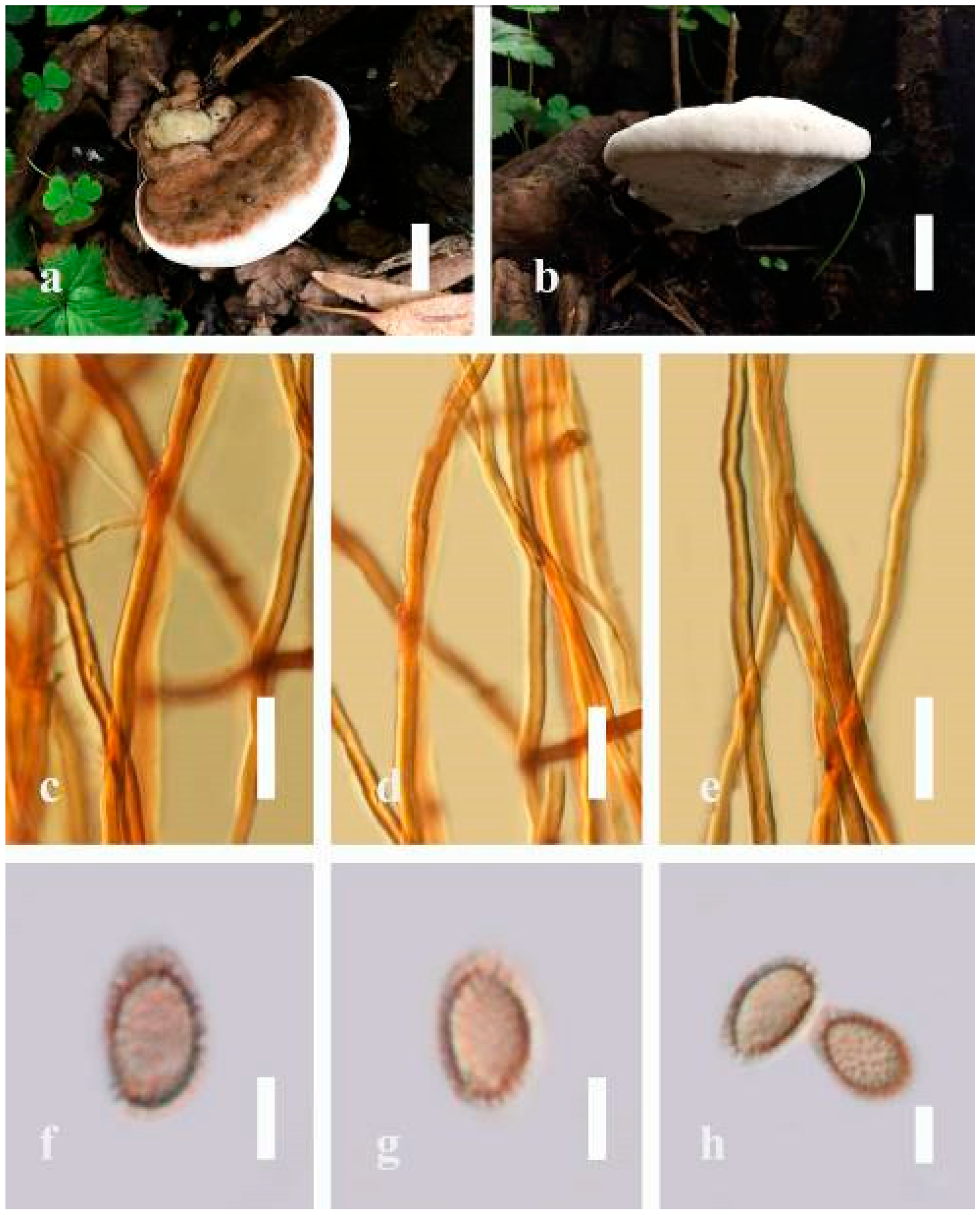{kind=link}
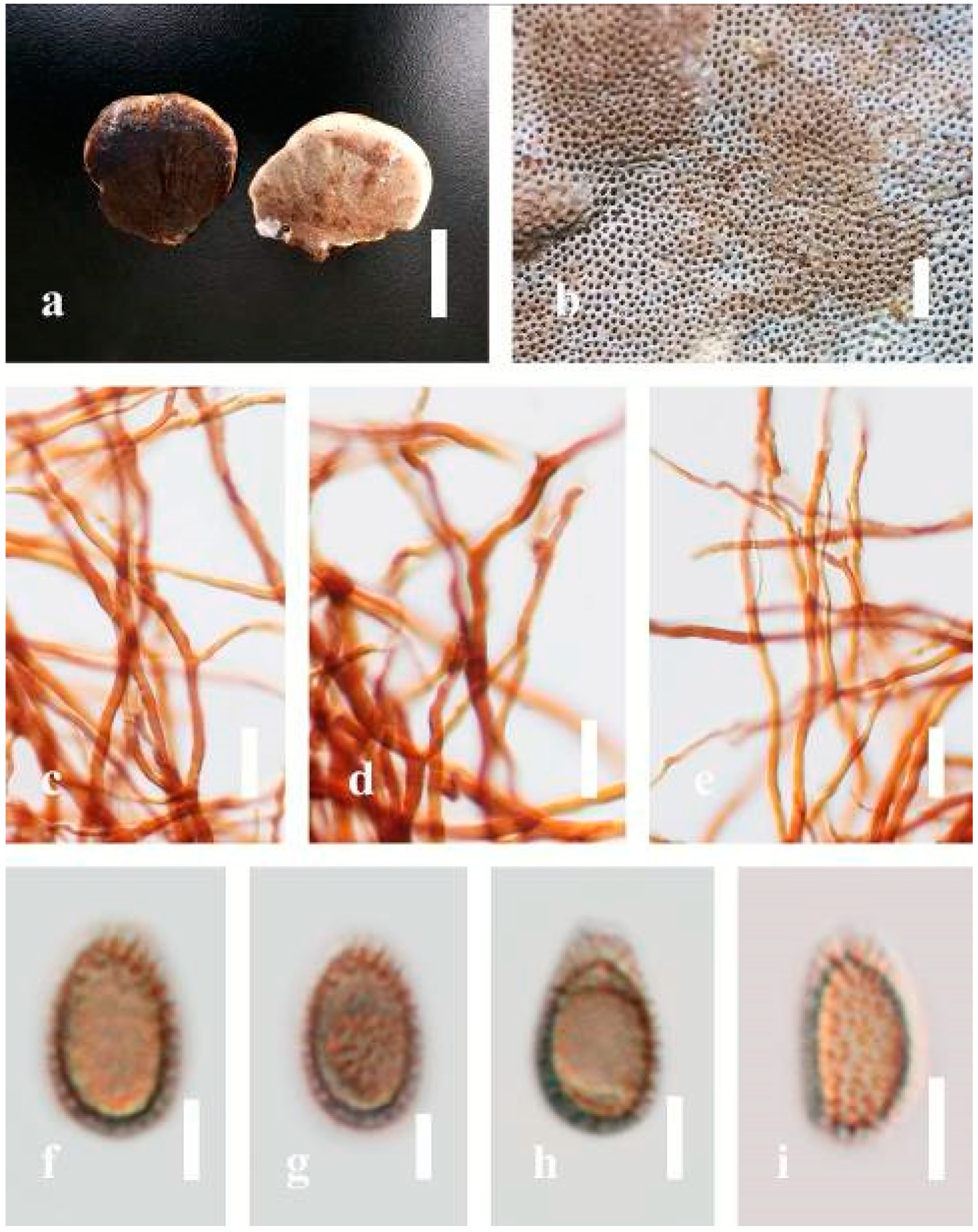{kind=link}
{kind=link}
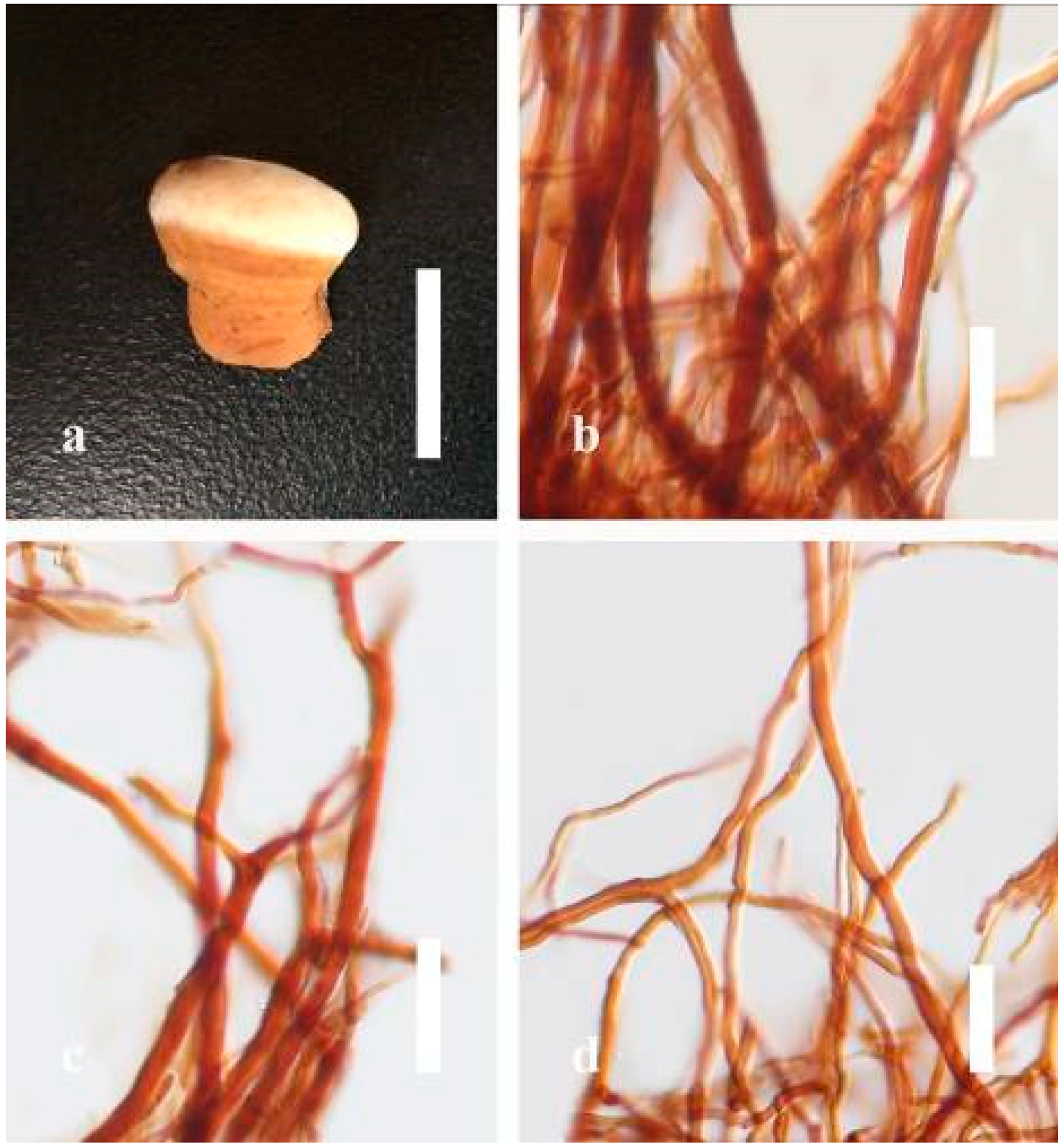{kind=link}
{kind=link}
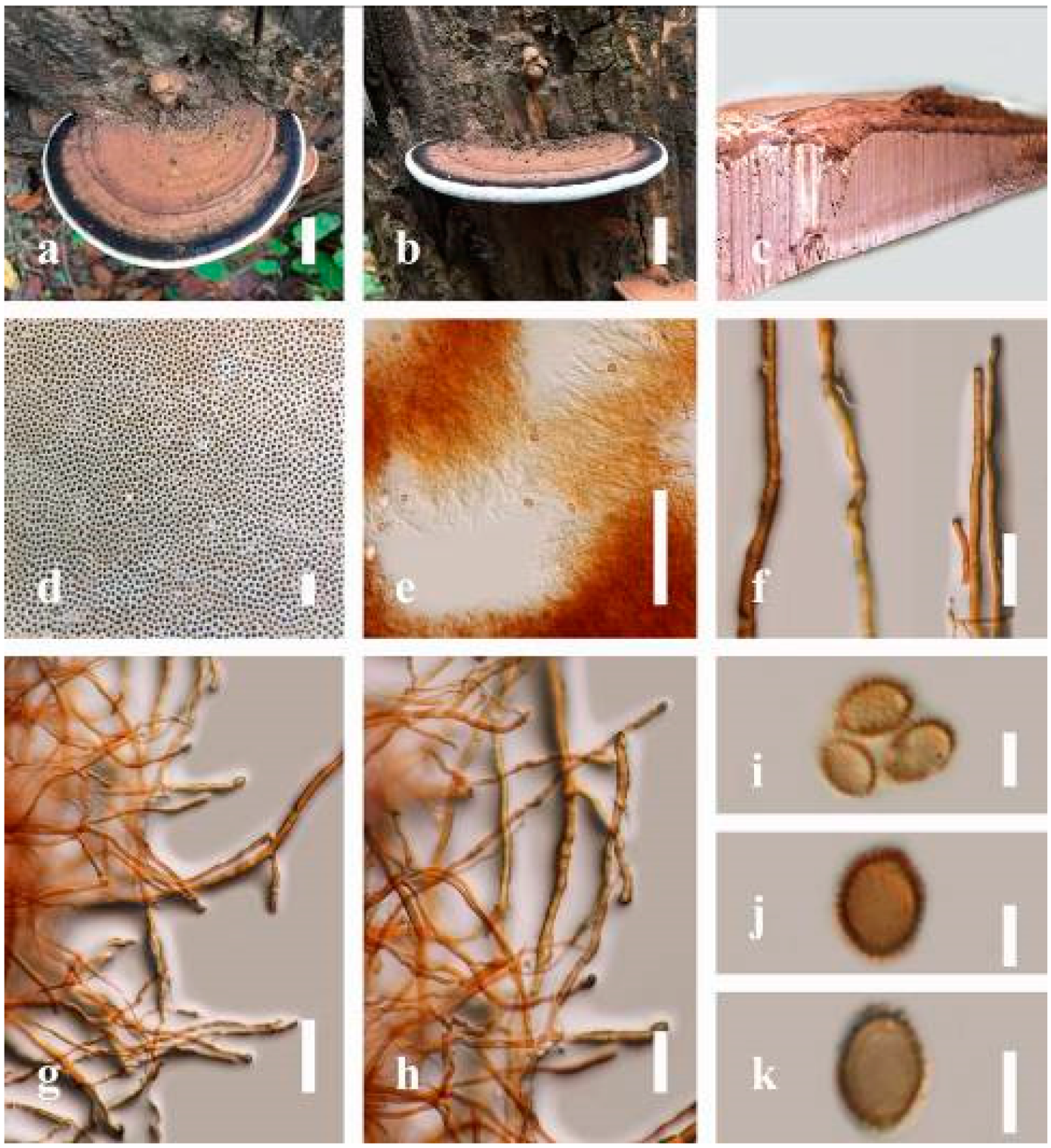{kind=link}
{kind=link}
{kind=link}
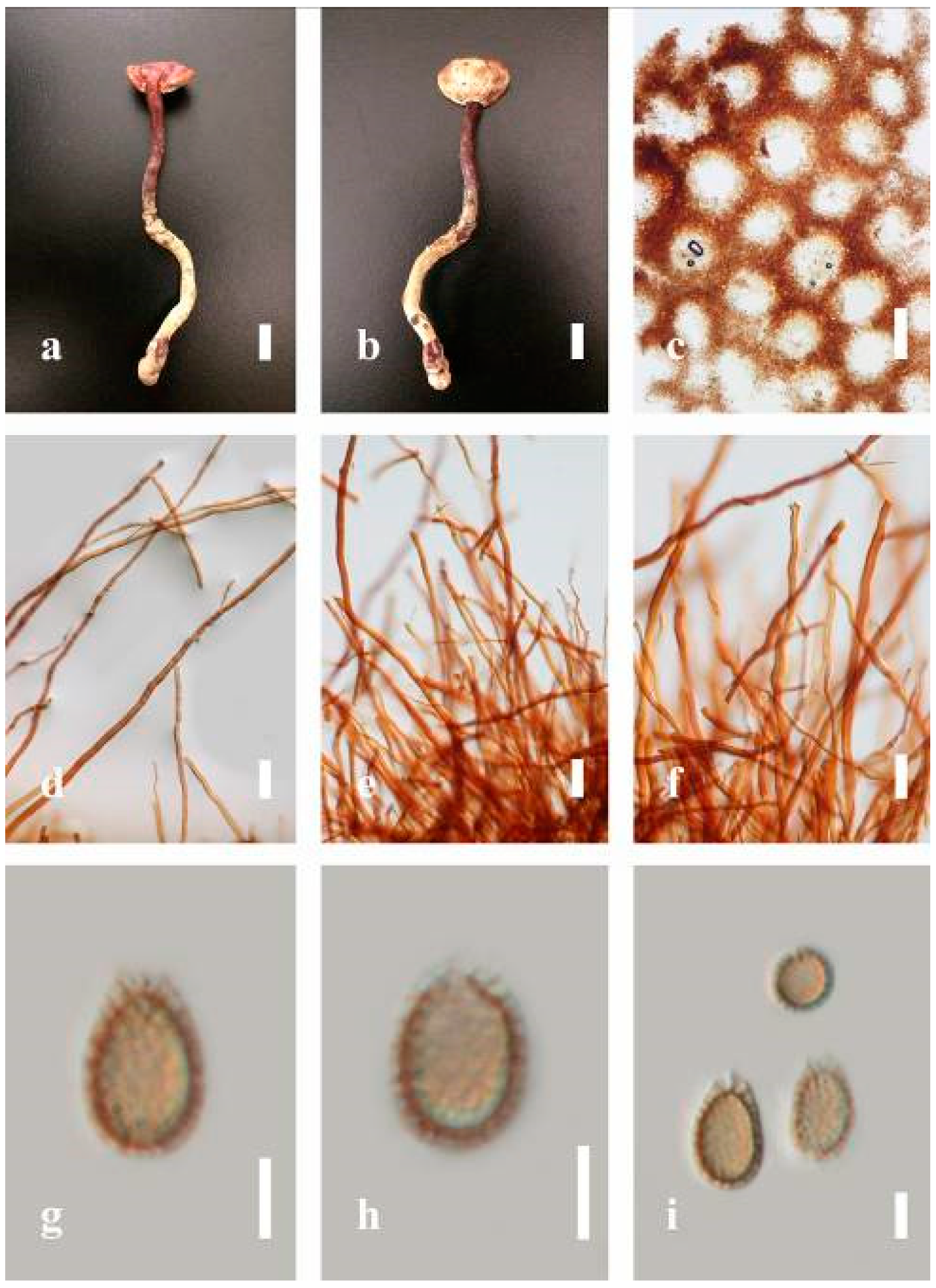{kind=link}
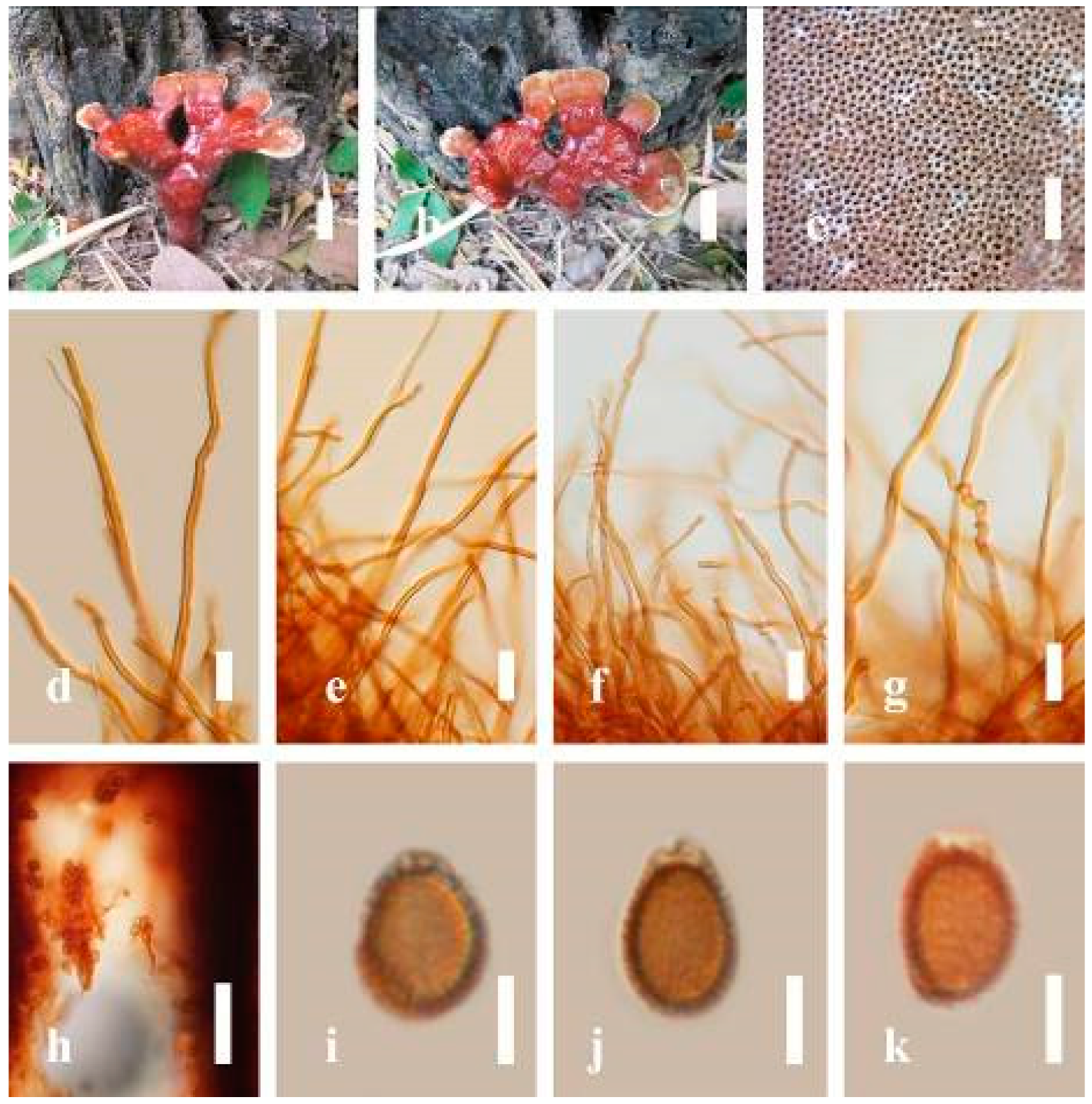{kind=link}
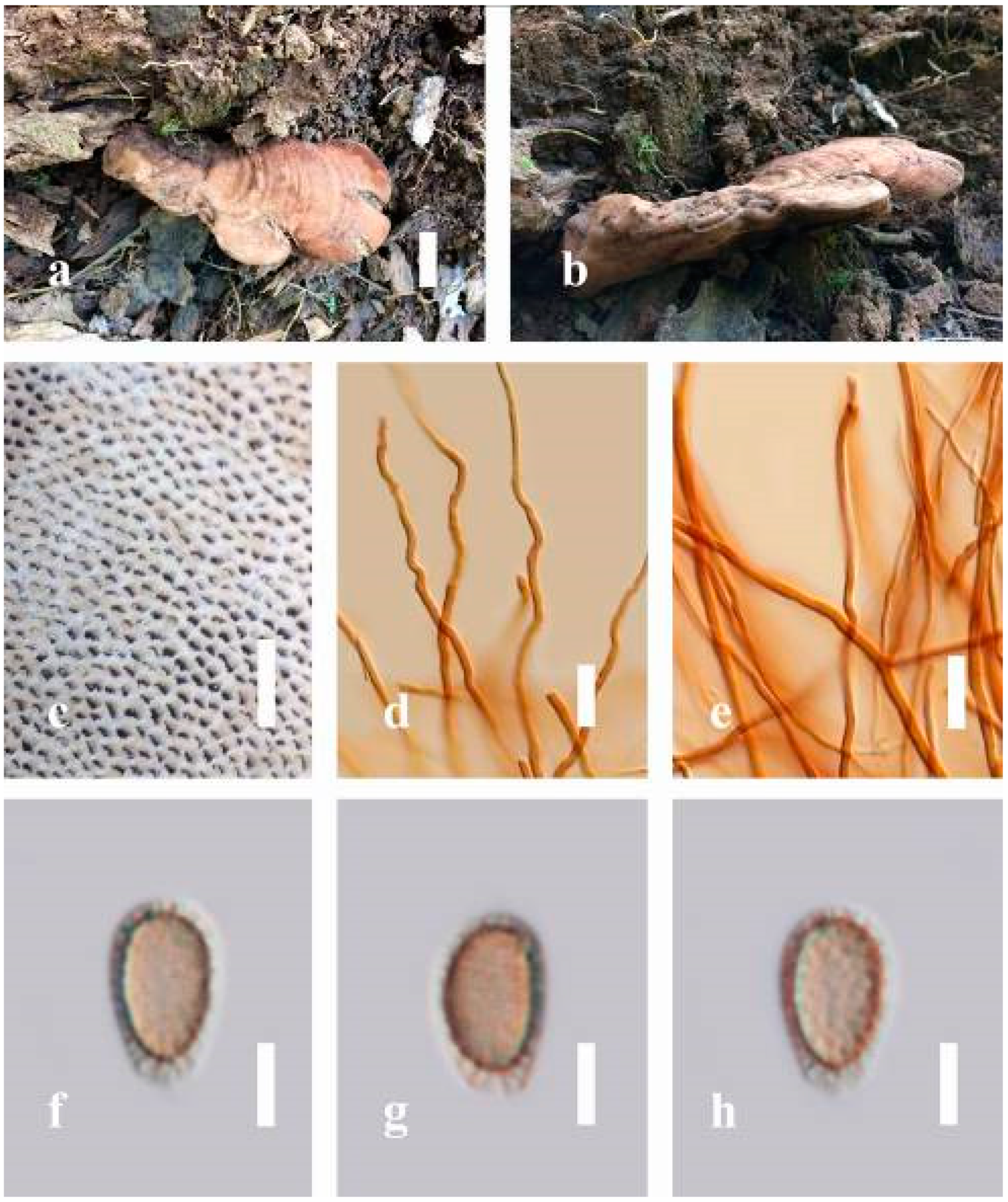{kind=link}
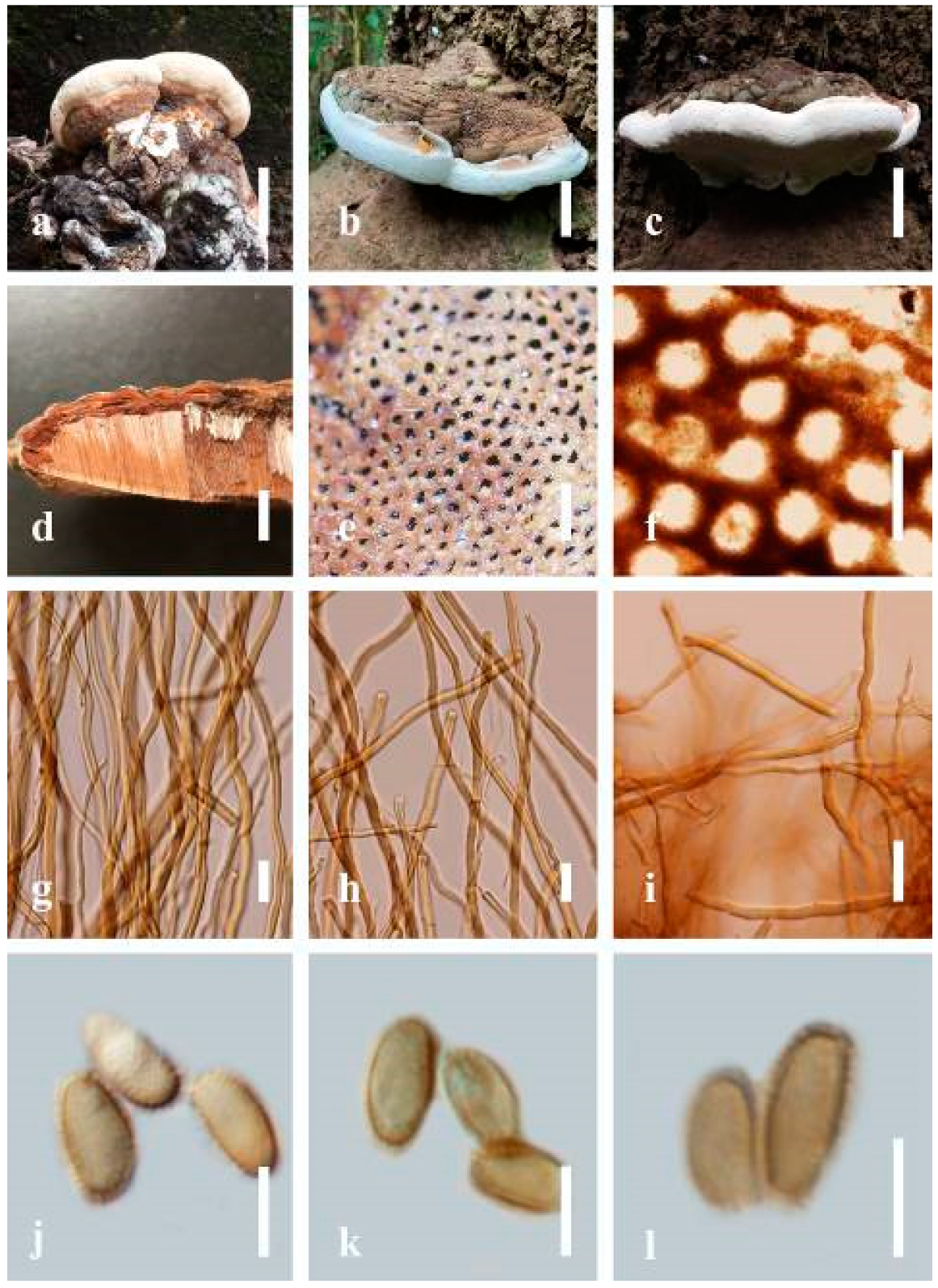{kind=link}
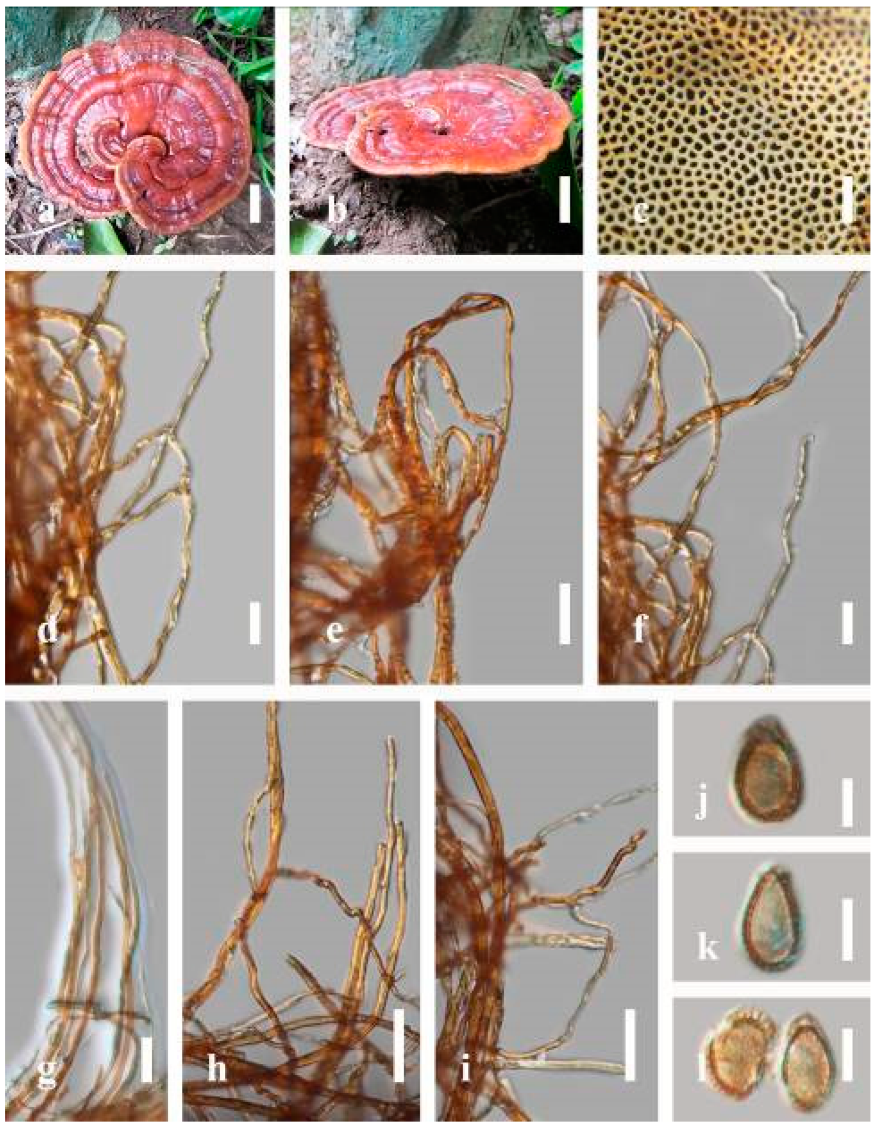{kind=link}
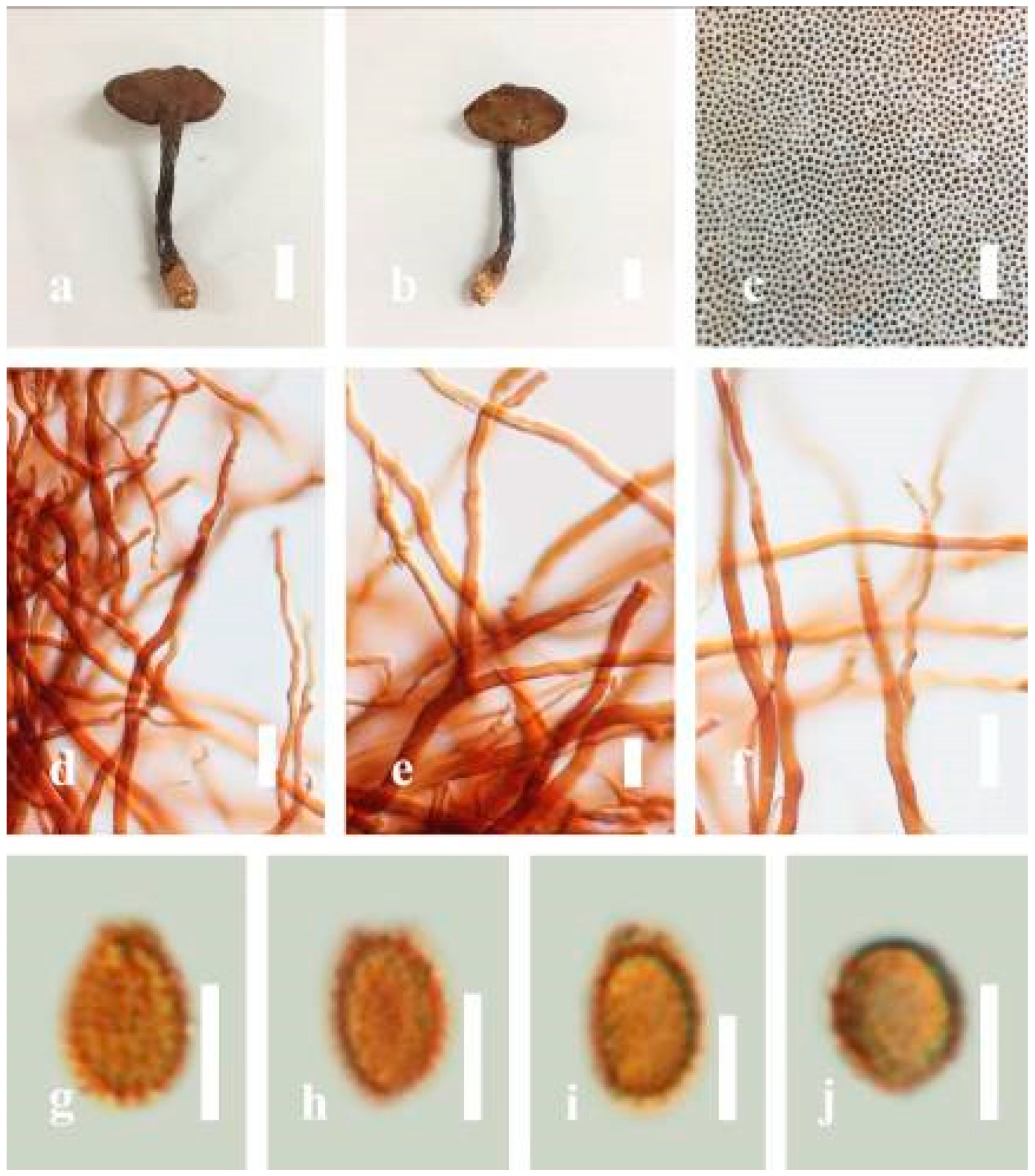{kind=link}
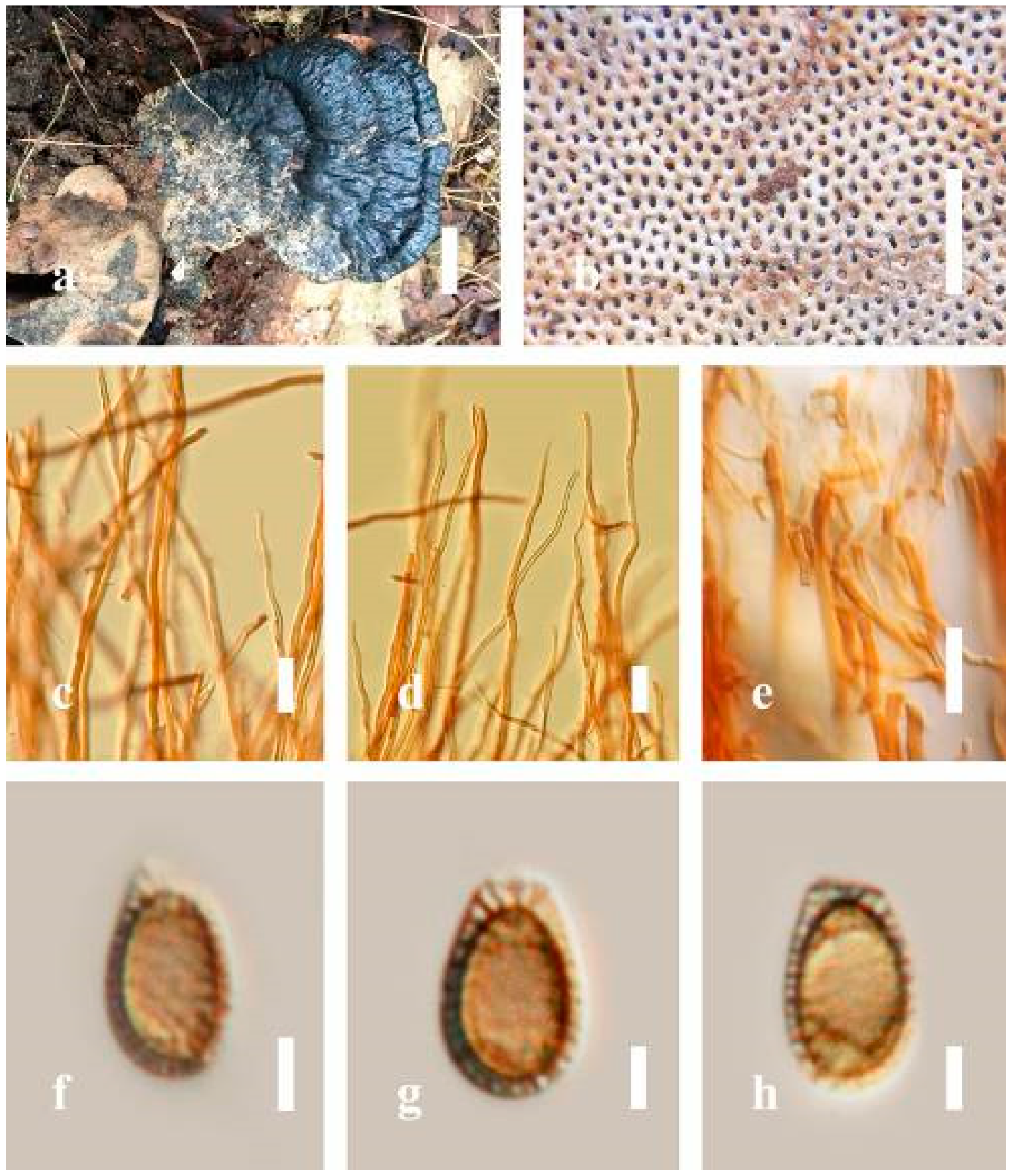{kind=link}
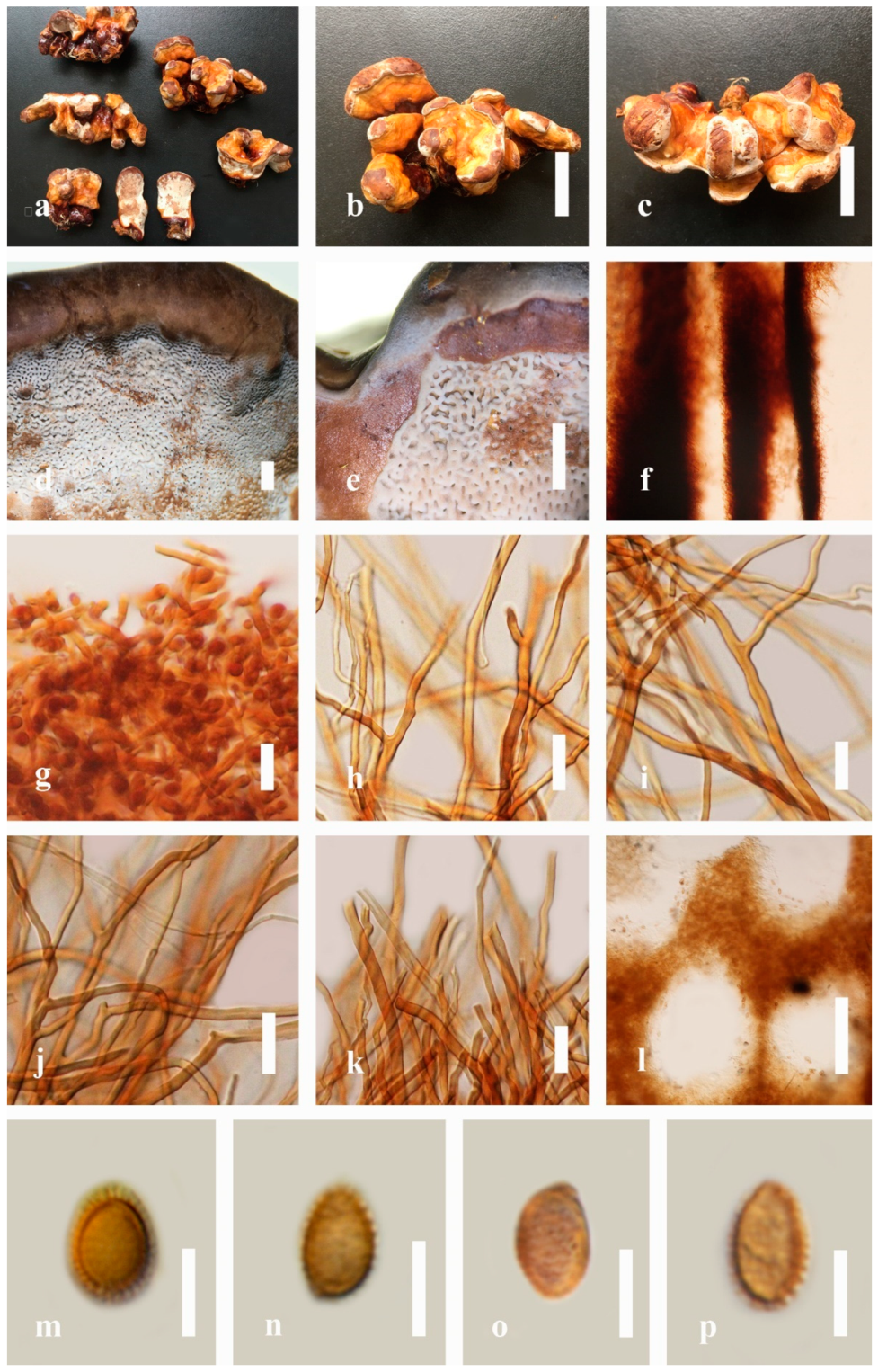{kind=link}


































